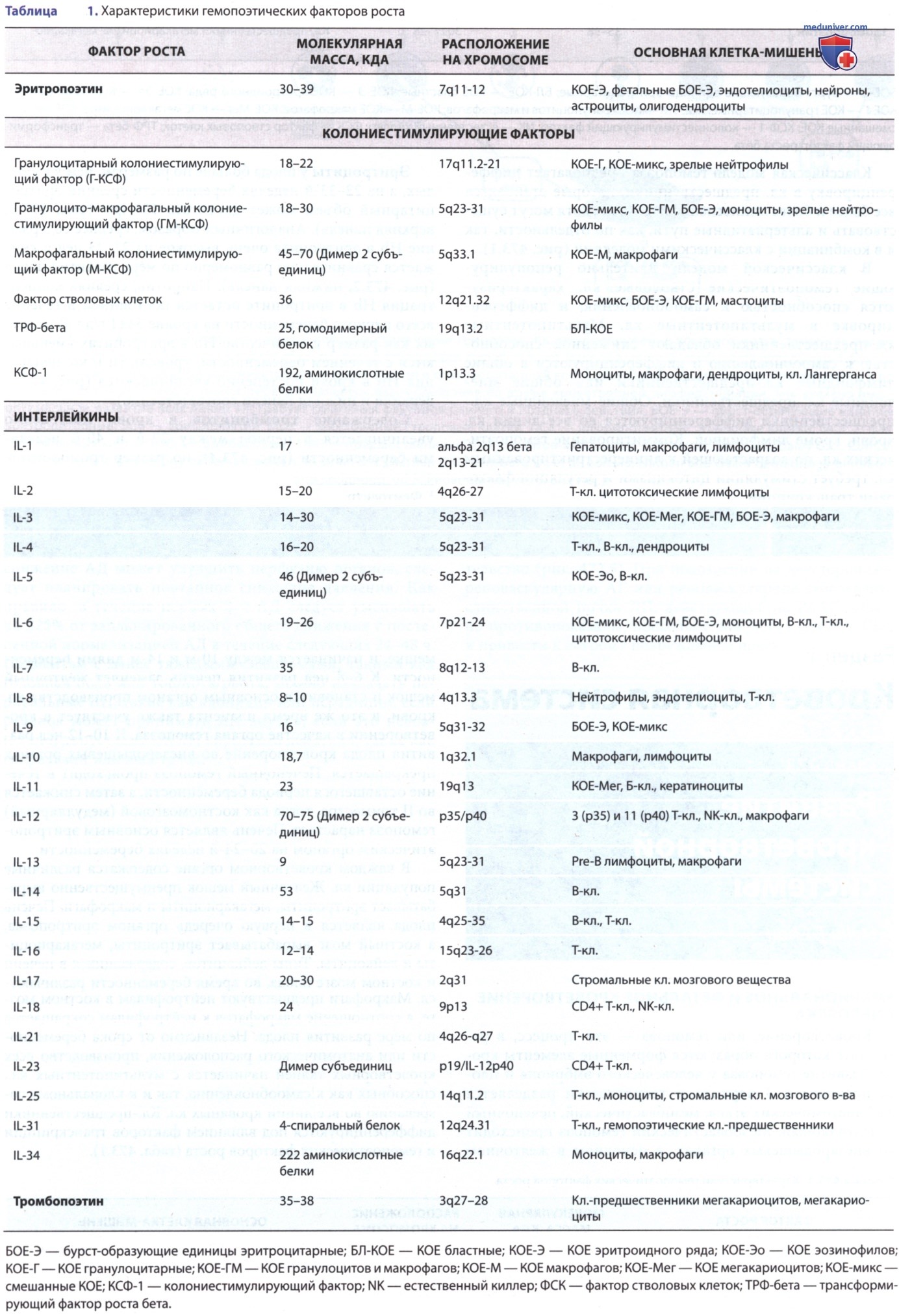
а) Эмбриональное и фетальное кроветворение у человека. Кроветворение, или гемопоэз — это процесс, в результате которого образуются форменные элементы крови. Развитие гемопоэза у человеческого эмбриона и плода проходит в 3 периода и теоретически разделяется на 3 анатомических этапа: мезобластический, печеночный и медуллярный. Мезобластический гемопоэз происходит во внезародышевых органах, в основном в желточном мешке, и начинается между 10-м и 14-м днями беременности. К 6-8 нед развития печень заменяет желточный мешок и становится основным органом производства кл. крови, в это же время плацента также участвует в кроветворении в качестве органа гемопоэза.
К 10-12 нед развития плода кроветворение во внезародышевых органах прекращается. Печеночный гемопоэз происходит в течение оставшегося периода беременности, а затем снижается во II триместре, тогда как костномозговой (медуллярный) гемопоэз нарастает. Печень является основным эритропоэтическим органом на 20-24-й неделях беременности.
В каждом кроветворном органе содержатся различные популяции кл. Желточный мешок преимущественно вырабатывает эритроциты, мегакариоциты и макрофаги. Печень плода является в первую очередь органом эритропоэза, а костный мозг вырабатывает эритроциты, мегакариоциты и лейкоциты. Типы лейкоцитов, содержащихся в печени и костном мозге плода, во время беременности различаются. Макрофаги предшествуют нейтрофилам в костном мозге, а соотношение макрофагов к нейтрофилам сокращается по мере развития плода. Независимо от срока беременности или анатомического расположения, производство всех кроветворных тканей начинается с мультипотентных кл., способных как к самообновлению, так и к клональному созреванию во все линии кровяных кл. Кл.-предшественники дифференцируются под влиянием факторов транскрипции и гемопоэтических факторов роста (табл. 1).
Классическая модель гемопоэза предполагает дифференцировку в кл.-предшественники, которые становятся все более и более линиеспецифическими, хотя могут существовать и альтернативные пути, как по отдельности, так и в комбинации с классическими моделями (рис. 1).
Рисунок 1. Основные источники цитокинов и способы стимуляции гемопоэза. Кл. микроокружения костного мозга, такие как макрофаги, кл. эндотелия и ретикулярные фибробласты, после стимуляции вырабатывают макрофагальный колониестимулирующий фактор, гранулоцито-макрофагальный колониестимулирующий фактор и гранулоцитарный колониестимулирующий фактор. Эти цитокины и др. кл., перечисленные в тексте, имеют перекрывающееся взаимодействие во время кроветворной дифференциации, как указано выше; оптимальное развитие всех линий кл. требует сочетания факторов раннего и позднего действия. БОЕ-Э — бурст-образующие единицы эритроцитарные; Эпо — эритропоэтин; МСК — миелоидные стволовые клетки; ПСК — плюрипотентные стволовые клетки; ФСК — фактор стволовых клеток; ТПО — тромбопоэтин; ГМП — предшественники гранулоцитов-моноцитов
В классической модели длительно репопулирующие гемопоэтические стволовые кл. характеризуются способностью к самообновлению и дифференцировке в мультипотентные кл. Мультипотентные кл.-предшественники обладают сниженной способностью к самообновлению и дифференцируются в общие лимфоидные кл.-предшественники или общие миелоидные кл.-предшественники. Общие миелоидные кл.-предшественники дифференцируются во все линии кл. крови, кроме лимфоидной. Коммитирование гемопоэтических кл. по возрастающей к линиерестриктированным кл. требует стимуляции цитокинами и регуляции факторами транскрипции.
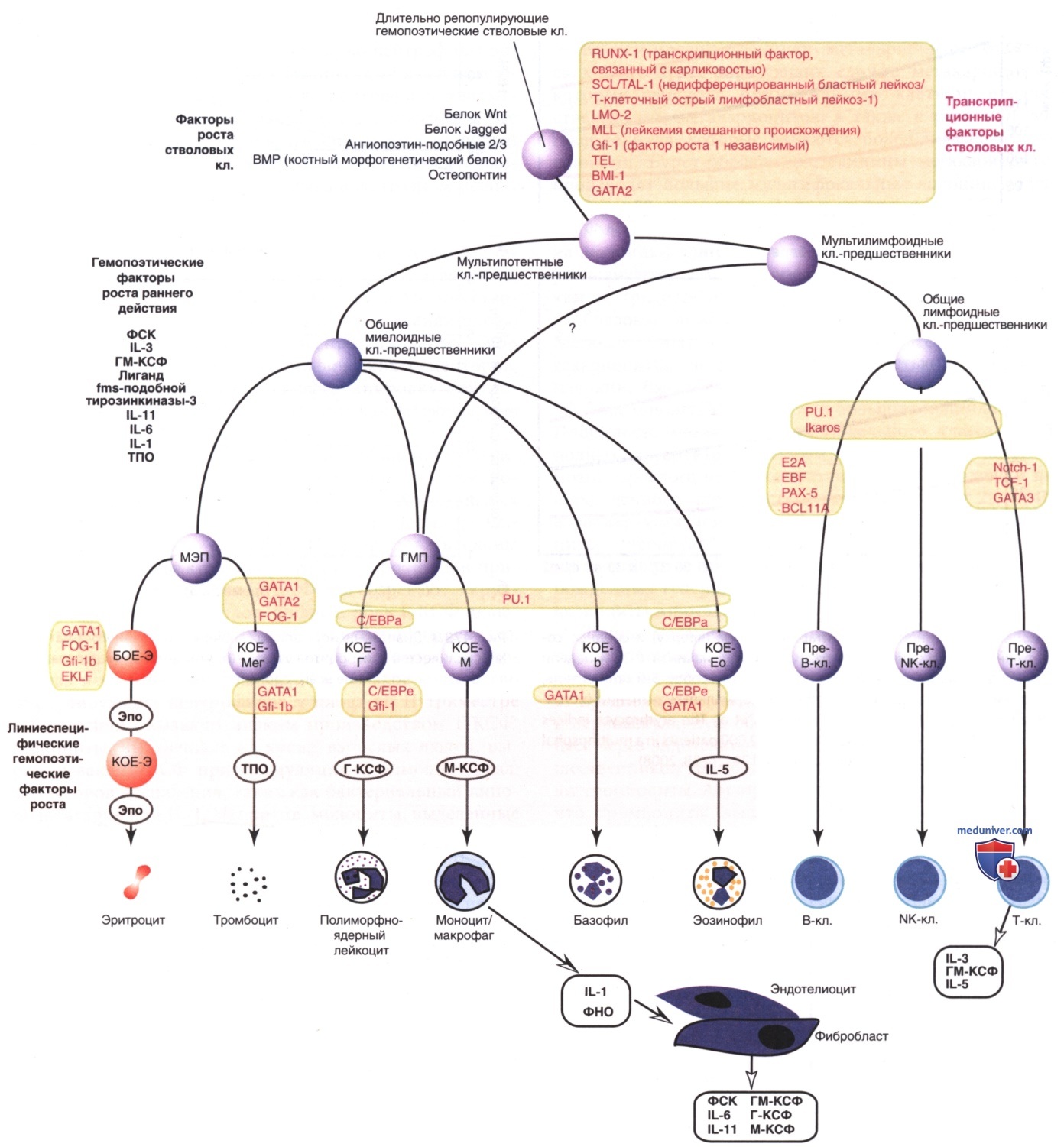
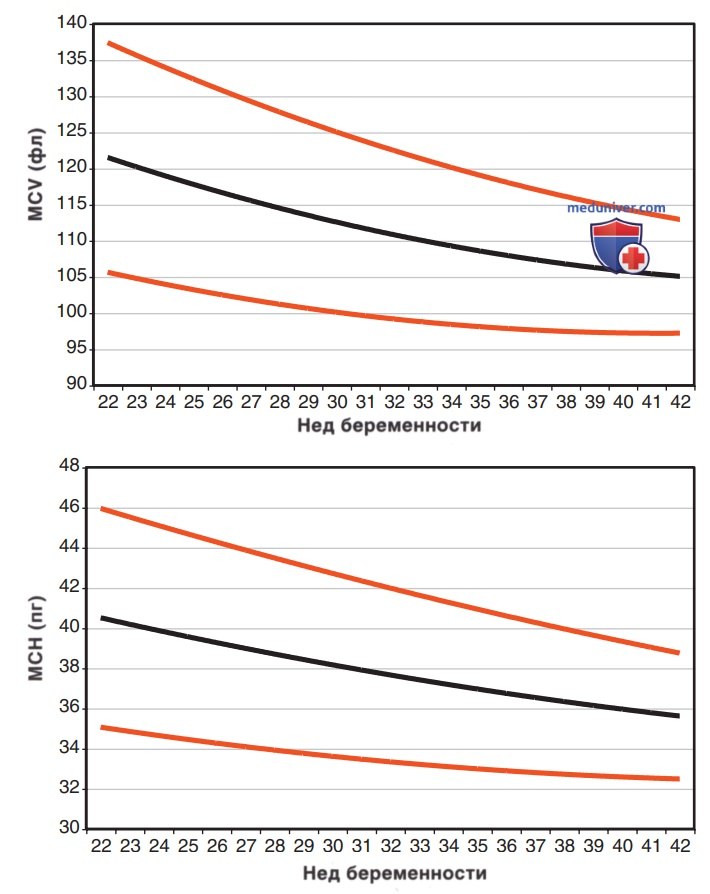
Эритроциты у плода больше по размерам, чем у взрослых, а на 22-23-й неделях беременности средний эритроцитарный объем может достигать 135 фл* (рис. 2, верхняя панель). Аналогичным образом, среднее содержание Hb в эритроците очень высокое на 22-23 нед и снижается сравнительно равномерно по мере развития плода (рис. 2, нижняя панель). Напротив, средняя концентрация Hb в эритроците остается постоянной в течение всего периода беременности на уровне 34± 1 г/дл. В то время как размер и количество Hb в эритроцитах уменьшаются с течением беременности, уровень Ht и концентрация Hb в крови постепенно увеличиваются (рис. 3, верхняя и нижняя панели соответственно).
P.S. * Фемтолитр.
Рисунок 2. Средний эритроцитарный объем (вверху) и среднее содержание гемоглобина в эритроците (внизу) начиная с 22-й недели до конца беременности. Линии представляют собой 5-й процентиль, медиану и 95-й процентиль диапазона нормальных значений
Рисунок 3. Диапазоны нормальных значений фетального гематокрита (А) и количества эритроцитов у плода (В) методом кордоцентеза в течение всего срока беременности
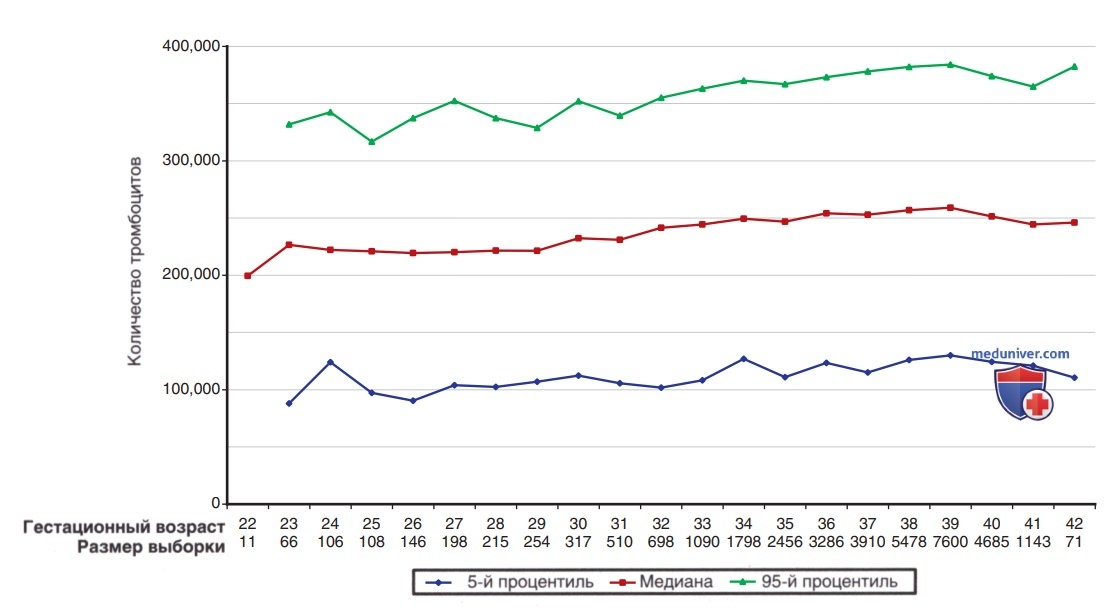
Содержание тромбоцитов в крови постепенно увеличивается в период между 22-й и 40-й неделями беременности (рис. 4), но размер тромбоцитов, оцениваемый по среднему объему тромбоцитов, остается постоянным на уровне 8±1 фл. Не наблюдается половых различий в диапазоне нормальных значений в фетальном и неонатальном периодах по отношению к эритроцитарным индексам, Ht, Hb, содержанию тромбоцитов или измерению среднего объема тромбоцитов.
Рисунок 4. Количество тромбоцитов начиная с 22-й недели и до конца беременности. Линии представляют собой 5-й процентиль, медиану и 95-й процентиль диапазона нормальных значений
б) Фетальный гранулоцитопоэз. Нейтрофилы впервые выявляются у эмбриона человека на ~5-й неделе после оплодотворения в виде небольших скоплений кл. вокруг аорты. Закладка костного мозга плода происходит на ~8-й неделе беременности, а на 8-10 нед объем костного мозга увеличивается, но нейтрофилы образуются там не ранее 10,5 нед. Начиная с 14 нед и на протяжении всего срока беременности нейтрофилы являются преобладающим типом гранулоцитов в костном мозге плода. Нейтрофилы и макрофаги образуются из общей кл.-предшественницы, но макрофаги появляются у плода раньше, чем нейтрофилы, сначала в желточном мешке, печени, легких и мозге, и все это происходит до того, как формируется костный мозг.
Г-КСФ и М-КСФ экспрессируются в костной ткани эмбриона уже на 6-й неделе после оплодотворения, а в печени плода — на 8-й нед. ГМ-КСФ и фактор стволовых кл. также хорошо представлены в тканях плода. Однако никакие изменения в экспрессии какого-либо из этих факторов или их специфических рецепторов, по-видимому, не отражают дифференцировку нейтрофилов или макрофагов у плода, так как маркеры еще не определены.
Кровь плода содержит мало нейтрофилов до II триместра. На 20 нед беременности количество нейтрофилов в крови составляет 0-500/мм3. Хотя зрелых нейтрофилов крайне мало, кровь плода изобилует кл,-предшественниками, способными генерировать клоны нейтрофилов. При культивировании в пробирке в присутствии рекомбинантного Г-КСФ они созревают в крупных колониях нейтрофилов. Физиол. роль Г-КСФ включает в себя регуляцию производства нейтрофилов, и это, по-видимому, относится как к плоду и новорожденному, так и к взрослому человеку. Т.о., низкое количество циркулирующих нейтрофилов у плода во II триместре м.б. частично вызвано низким производством Г-КСФ.
Моноциты, выделенные из крови взрослых людей, вырабатывают Г-КСФ при стимуляции с помощью разл. медиаторов воспаления, таких как бактериальный липополисахарид или IL-1. Напротив, моноциты, выделенные из крови или органов плода до 24-й недели беременности, после стимуляции липополисахаридов или IL-1 генерируют лишь небольшое количество Г-КСФ белка и матричной РНК. Тем не менее рецепторы Г-КСФ на поверхности нейтрофилов новорожденных равны по количеству и подобны тем, которые имеются на нейтрофилах у взрослых.
Функция гранулоцитопоэтических факторов (Г-КСФ, М-КСФ, ГМ-КСФ, фактор стволовых кл.) у плода не ограничивается кроветворным процессом. Рецепторы для каждого из этих факторов расположены в областях ЦНС и ЖКТ плода, где с развитием плода изменяется их профиль экспрессии.
в) Фетальный тромбопоэз. Между мегакариопоэзом и тромбопоэзом у плода/новорожденного и у взрослого человека присутствует несколько биологических различий. Существует уникальный возрастной паттерн фетального/неонатального мегакариопоэза, характеризующийся быстрой пролиферацией с последующим полным цитоплазматическим созреванием без полиплоидизации. Фетальные и неонатальные мегакариоциты значительно меньше по размеру, имеют более низкую плоидность и производят меньшее количество тромбоцитов. Однако у фетальных и неонатальных мегакариоцитов пролиферативный потенциал выше, чем у кл.-предшественников взрослого человека.
Эти различия позволяют быстрорастущему плоду и новорожденному заполнять кл. костный мозг и кровь, сохраняя при этом нормальное количество тромбоцитов.
Клетки-предшественники мегакариоцитов относятся к ряду бурст-образующих единиц мегакариоцитов, которые являются примитивными клетками-предшественниками мегакариоцитов, а также к ряду КОЕ мегакариоцитов, которые являются более дифференцированными. Бурст-образующие единицы мегакариоцитов производят большие мультифокальные колонии, содержащие >50 мегакариоцитов, в то время как КОЕ мегакариоцитов производят меньшие по размеру (3-50 кл. на колонию) унифокальные колонии. Мегакариоциты распознаются по их морфологическим характеристикам в процессе эндоредупликации, которая приводит к образованию крупных кл. с полиплоидными ядрами. Мегакариоциты, в отличие от кл.-предшественников мегакариоцитов, не обладают способностью генерировать колонии.
Вместо этого они созревают, переходя от маленьких мононуклеарных кл. к большим полиплоидным. Плоидность мегакариоцитов модального класса (число полных комплектов хромосом) у нормального костного мозга взрослого человека составляет 16N. У плода и новорожденного плоидность ниже, в основном 2N и 4N, а размер мегакариоцитов меньше. Большие мегакариоциты генерируют больше тромбоцитов, чем маленькие мегакариоциты; предполагается, что мегакариоциты новорожденных генерируют меньше тромбоцитов, чем мегакариоциты взрослых людей.
Точные механизмы, с помощью которых мегакариоциты высвобождают тромбоциты в циркуляцию, до сих пор до конца не изучены. Изучение этого процесса позволяет предположить, что зрелые мегакариоциты мигрируют в периваскулярную область и распространяют этот процесс через эндотелий, что приводит к образованию предшественников тромбоцитов, которые затем высвобождают тромбоциты. Альтернативный путь заключается в том, что тромбоциты высвобождаются из мегакариоцитов в легких в результате сил сдвига.
Тромбопоэтин (ТПО) является основным регулятором развития мегакариоцитов и производства тромбоцитов (см. табл. 1). Тромбороэтин преимущественно вырабатывается в печени с самого начала фетального периода до взрослой жизни, но также экспрессируется кл. в почках и в меньшей степени кл. гладкой мускулатуры и костного мозга. У здоровых новорожденных любого гестационного возраста концентрация тромбороэтина выше, чем у здоровых взрослых. ТПО является основным стимулятором производства мегакариоцитов и тромбоцитов, но фактор стволовых кл., IL-3, IL-11, IL-6 и эритропоэтин также стимулируют мегакариопоэз и тромбоэз in vitro и in vivo.
Важно, что тромбороэтин также способствует экспансии гемопоэтических стволовых кл. и кл.-предшественников, а рецептор тромбороэтина экспрессируется на гемопоэтические столовые кл. и эритроидных предшественниках в дополнение к предшественникам мегакариоцитов, мегакариоцитов и зрелых тромбоцитов.
г) Фетальный эритропоэз. Подобно гемопоэзу других клеточных линий, фетальный эритропоэз регулируется факторами роста, продуцируемыми плодом, а не организмом матери. Эритропоэтин не проникает через плаценту человека. Стимуляция производства материнского эритропоэтина не увеличивает эритропоэз плода, а подавление материнского эритропоэза путем гипертрансфузии не подавляет эритропоэз плода. Эритропоэтин играет ключевую роль в регуляции пролиферации и созревания эритроидных прешественников. Эритроидные коммитированные кл.-предшественники состоят из бурст-образующих единиц эритроцитарного ряда и КОЕ эритроцитарного ряда.
При количественном определении КОЕ, бурст-образующие единицы эритроцитарные человека более пролиферативные, они образуют колонии из множества кластеров эритробластов по контрасту с КОЕ эритроцитарного ряда, которые образуют 1 или 2 кластера, каждый из которых содержит 8-100 ге-моглобинизированных эритробластов.
Эритропоэтин необходим для производства эритроцитов из кл. КОЕ эритроцитарного ряда путем стимуляции выживания и пролиферации эритробластов. Эритропоэтин связывается со специфическими рецепторами на поверхности эритроидных коммитированных кл.-предшественников, и его экспрессия регулируется чувствительным к кислороду механизмом через семейство белков гипоксия-индуцируемого фактора (HIF; англ. hypoxia-inducible factor). HIF-1α и HIF-2α регулируются парциальным давлением кислорода, тогда как HIF-1β конститутивно экспрессируется. Вместе белки HIF поддерживают кислородный гомеостаз и регулируют эритропоэз, индуцируя эритропоэтина в условиях гипоксии.
Эритропоэтин вырабатывается моноцитами и макрофагами в печени плода в течение I и II триместров. После рождения анатомическая область производства эритропоэтина смещается в почку. Специфический стимул такого сдвига неизвестен, но м.б. связан с повышением давления артериального кислорода, которое имеет место при рождении. Эпигенетическая регуляция экспрессии генов также может играть определенную роль, так как оказывается, что почечные и печеночные гены эритропоэтина в разной степени метилированы. Хотя матричная РНК эритропоэтина и белок м.б. обнаружены в почке плода, неизвестно, является ли такой способ производства биологически существенным.
По-видимому, производство эритропоэтина почками не является необходимым для нормального фетального эритропоэза, о чем свидетельствуют нормальные значения концентрации эритропоэтина в сыворотке и Ht у плодов с отсутствующей почкой.
1. Гемоглобин у плода и новорожденного. Гемоглобин — это тетрамер из 4 цепей глобинов с железосодержащим порфириновым кольцом, называемым гемом, связанным с каждой цепью
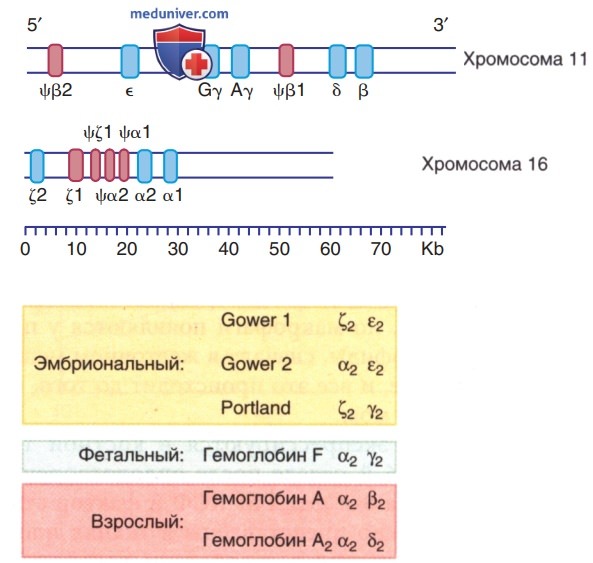
Динамическое взаимодействие гема и глобина придает Hb уникальные свойства при обратимом транспорте кислорода. Молекула Hb состоит из 2α-подобных и 2β-подобных полипептидных цепей, к каждой из которых прикреплена гемовая группа. Кластеры генов α-глобина и β-глобина расположены на хромосоме 16 и 11 соответственно (рис. 5). Существует 2β-глобиновых гена и 4 a-глобиновых гена. В эритроцитах эмбриона, плода, ребенка и взрослого обычно м.б. обнаружены 6 различных Hb (рис. 6): эмбриональные Hb (Говер-1, Говер-2 и Портленд), фетальный НЬ (HbF) и Hb взрослых людей (HbА и HbА2). Электрофоретическая подвижность Hb варьируется в зависимости от их хим. структуры.
Рисунок 5. Организация глобиновых генов. Нижняя линия отражает шкалу в килобазах. Верхний сегмент представляет β-подобные глобиновые гены на хромосоме 11, а нижний сегмент — α-подобные гены на хромосоме 16. Области гена, кодирующие первичные белки глобина, показаны в виде синих сегментов, а области, кодирующие псевдогены («ф», невыраженные остатки), показаны в виде розовых сегментов. Приведен состав эмбрионального, фетального и взрослого Hb. α; β; γ; δ; ε; ζ
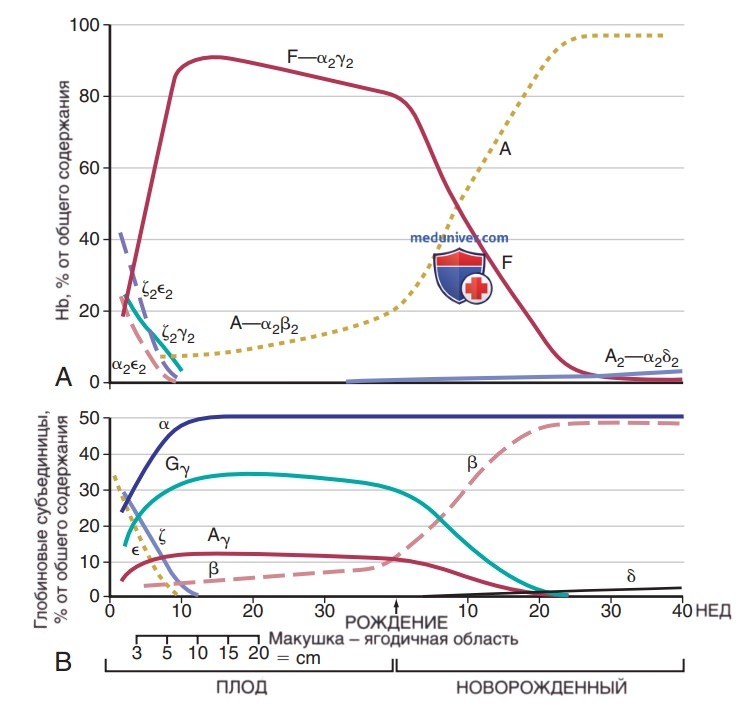
Рисунок 6. Изменения в тетрамерах гемоглобина (А) и глобиновых субъединицах (В) в процессе развития человека от эмбриона до раннего младенчества
Экспрессия и количественные соотношения Hb определяются сложными процессами развития. Экспрессия глобиновой цепи является специфической стадией развития и происходит через 2 гемоглобиновых перехода, медиируемых в первую очередь изменениями экспрессированных генов β-глобина. Существует 5 функциональных β-подобных генов глобиновой цепи: эмбриональный (НВЕ1), 2 фетальных (HBG1, HBG2) и 2 взрослых (HBD, НВВ); и 3α-подобных гена глобиновой цепи: эмбриональный (HBZ) и 2 взрослых (НВА1, НВА2). Примитивные эритроидные кл. главным образом экспрессируют эмбриональные глобины. 1-й переход β-глобина возникает ~ на 6-й неделе беременности в фетальный глобин (HbG), что совпадает с началом дефинитивной стадии развития гемопоэза. Основной HbF состоит из 2α- и 2γ- глобиновых цепей (а2γ2).
Второй глобиновый переход отвечает за экспрессию основного Hb нормального взрослого человека (HbА), состоящего из 2α- и 2β-полипептидных цепей (а2β2), и впервые экспрессируется в середине беременности. Ключевым регулятором перехода HbF в Hb взрослого человека является транскрипционный фактор BCL11А, который связывается с геном β-глобина и действует на подавление экспрессии γ-глобина и, т.о., экспрессии HbF.
2. Эмбриональные типы гемоглобинов. Кровь эмбрионов человека на ранних стадиях развития содержит 2 медленно мигрирующих типа Hb — Говер-1 и Говер-2, а также НЬ Портленд, который обладает HbF-подобной подвижностью. Дзета (ζ) цепи Hb Портленд и Говер-1 структурно похожи на a-цепи. Оба типа Hb Говер содержат эпсилон (ε) β-подобную полипептидную цепь глобина. Hb Говер-1 имеет структуру (2е2, в то время как Говер-2 имеет структуру а2е2. НЬ Портленд имеет структуру ζ2γ2. В эмбрионах до 6-й недели беременности преобладают типы Hb Говер, но к 3 мес беременности они больше не обнаруживаются.
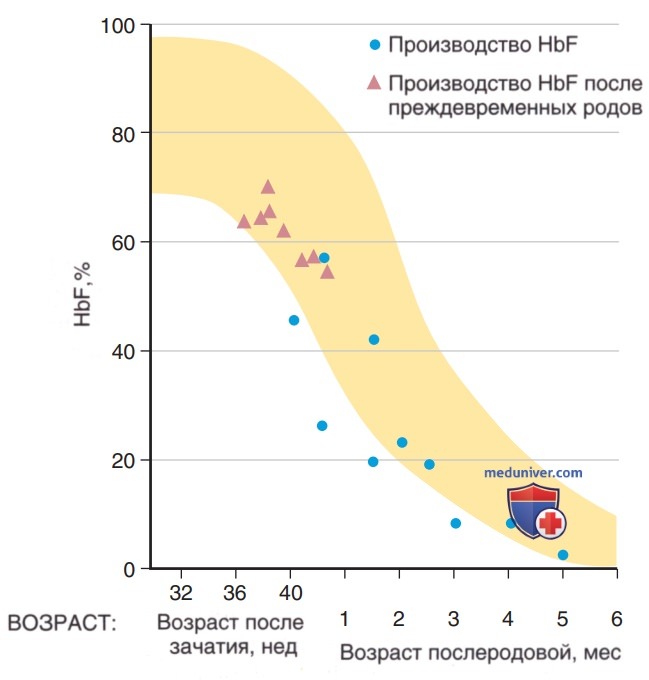
3. Фетальный гемоглобин. К 6-8-й неделям беременности HbF (а2у2) является доминирующей формой Hb, а к 24-й неделе беременности он составляет 90% от общего Hb. Уровень HbF умеренно снижается в III триместре и составляет 70-80% от общего объема Hb. Образование HbF быстро снижается в постнатальном периоде (рис. 7) и к 6-12 мес достигает уровня <2% от взрослого возраста. Понимание молекулярной основы перехода HbF в Hb взрослого человека представляет интерес из-за терапевтической пользы для пациентов с β-талассемией и СКА, клиническая тяжесть которых улучшается при умеренном ↑ уровня HbF. Точные механизмы, с помощью которых BCL11A угнетает HbF, не полностью изучены, но определены эритроидные специфические усилители BCL11А, которые являются потенциальными мишенями для терапевтической HbF-индукции.
Рисунок 7. До- и послеродовые изменения в % от общего гемоглобина, представленного фетальным гемоглобином (желтый цвет). Треугольники представляют постнатальное производство ретикулоцитов у недоношенных младенцев, а круги — пуповинную кровь и постнатальное производство ретикулоцитов у доношенных детей
4. Гемоглобин у взрослых людей. Уровень HbА составляет 5-10% от общего Hb на 24-й неделе беременности и продолжает постепенно увеличиваться, так что в среднем HbА составляет 30% от общего Hb на протяжении всего срока беременности. К 6-12 мес дети достигают уровня НЬА как у взрослого человека. Второстепенный зрелый гемоглобин — НЬА2 — содержит δ цепи и имеет структуру а.282. При рождении уровень НЬА2 составляет <1,0%, но к 12 мес нормальный уровень составляет 2,0-3,4%. На протяжении всей жизни нормальное соотношение НЬА к НЬА2 составляет 30:1.
5. Изменения уровня гемоглобина из-за болезней. Уровень HbF может повышаться при гемоглобинопатии, наследственном персистировании HbF или синдромах недостаточности костного мозга, а также м.б. связан с эритропоэзом при стрессовых ситуациях. Поскольку уровень HbF ↑ в течение первого года жизни, важно знать его нормальную картину снижения (см. рис. 6 и 7). Два заболевания, возникающие в результате мутаций в гене β-глобина (НВВ), β-талассемия и СКА, становятся симптоматичными в постнатальном периоде по мере снижения экспрессии γ-глобина у плода и увеличения экспрессии взрослого β-глобина. При обоих этих заболеваниях, повышенный уровень HbF сохраняется в детском возрасте и у взрослых.
У пациентов с наиболее тяжелым видом заболевания, β0-талассемией, HbF является единственным производимым типом Hb, за исключением небольшого количества HbА2. На другом конце спектра у пациентов с малой β-талассемией послеродовое ↓ HbF замедляется, и слегка повышенные уровни HbF (>2%) могут сохраняться на протяжении всей жизни.
Пациенты с СКА, которые также имеют мутацию в гене НВВ, как правило, демонстрируют повышенные уровни HbF, варьирующие от 5 до 30%.
Напротив, повышенный уровень HbF не характерен для синдромов α-талассемии, но в неонатальном периоде м.б. обнаружены тетрамеры γ-цепей (γ4 или Hb Barts). Так как α-глобиновые цепи экспрессируются в Hb плода и взрослого, то 4-α-генные мутации, приводящие к функциональным делециям, не совместимы с жизнью. Плоды погибают в утробе или вскоре после рождения от тяжелой анемии и отечного синдрома новорожденных. Наследование только 1 нормального гена 4 (α-/-) приводит к возникновению Н-гемоглобинопатии, которая обычно ассоциируется с анемией средней степени тяжести. Наследование 2 или 3 нормальных a-генов приводит к появлению малой а-талассемии или статуса носителя, соответственно.
Наследственное персистирование фетального гемоглобина — доброкачественное генетическое заболевание, вызванное гетерозиготными делециями или замещениями нуклеотидов в областях β-глобинового локуса, которые регламентируют транскрипцию генов HBG1 и HBG2, вызывая персистирующие уровни экспрессии HbF во всех кл., составляющие около 30% от общего количества гемоглобинов. Пациенты с наследственнымм персистирование фетального гемоглобина не страдают анемией.
У недоношенных младенцев, получавших рекомбинантный эритропоэтин человека, увеличивается выработка HbF во время активного эритропоэза. Умеренное повышение уровня HbF может также встречаться при многих заболеваниях, сопровождающихся гематологическим стрессом, таких как гемолитическая анемия, лейкемия и синдромы недостаточности костного мозга, напр. анемия Даймонда-Блэкфана (Blackfan, Kenneth. Diamond, Louis).
Нормальный уровень HbА2 у взрослого человека (2,0-3,4%) изменяется редко. Уровни HbА2 >3,4% встречаются у большинства людей с малой β-талассемией, а также у людей с мегалобластическими анемиями, вторичными по отношению к дефициту витамина В12 и фолиевой кислоты. Снижение уровней HbА2 наблюдается при ЖДА и α-талассемии.
д) Продолжительность жизни эритроцитов у плода и новорожденного. Как правило, самый высокий Ht в течение жизни человека наблюдается при рождении, а самый низкий Ht — при физиол. минимуме, который возникает на 8-10 нед жизни. Укороченная продолжительность жизни фетальных и неонатальных эритроцитов предположительно является важным фактором. Средняя продолжительность жизни эритроцитов у нормальных взрослых составляет 120 дней.
Срок жизни фетальных/неонатальных эритроцитов когда-то оценивался значительно меньше — в среднем 60-90 дней, как показывали исследования эритроцитов, меченных хромом (51Cr). Однако новые исследования показывают, что продолжительность жизни фетальных/неонатальных эритоцитов аналогична продолжительности жизни эритроцитов взрослого человека. Неоцитолиз — это активное удаление молодых эритроцитов, которые были сформированы в условиях относительной гипоксии после нормо- или гипероксических состояний. Этот процесс был также предложен в качестве объяснения физиол. минимума новорожденных.
Видео лекция кровь как ткань, кроветворение, лимфа