Стероидные гормоны и пептидные факторы различаются по своему специфическому рецепторно-опосредованному воздействию. Однако и те и др. в конечном счете приводят к транскрипции ДНК и производству белка в клетке-мишени.
а) Сопряженные с G-белком рецепторы. Это мембранные рецепторы, которые связывают пептидные факторы. Рецепторы состоят из гидрофильного внеклеточного, в/клеточного и гидрофобного трансмембранного доменов, которые по спирали пересекают клеточную мембрану 7 раз.
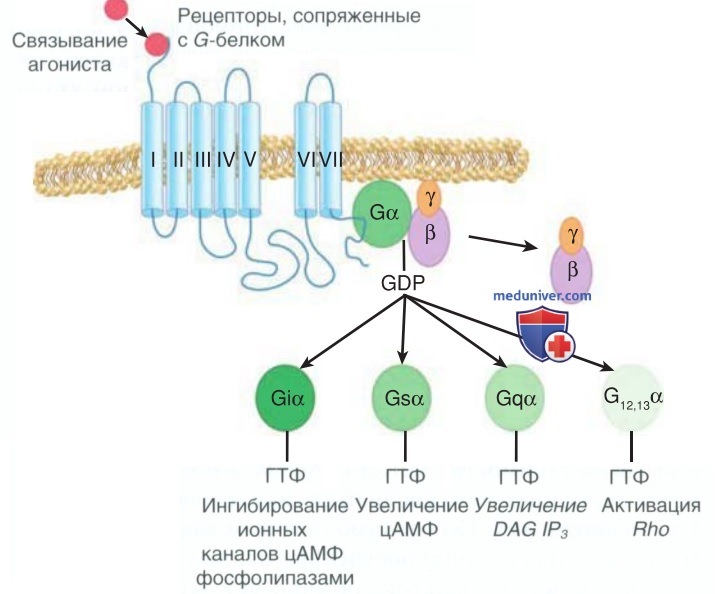
При связывании с гормоном данные рецепторы претерпевают конформационные изменения, активируют в/клеточные сигнальные пути и вследствие фосфорилирования изменяют транскрипцию нескольких генов в клетке-мишени (рис. 1).
Рисунок 1. Рецепторы, сопряженные с G-белком, представляют собой интегральные трансмембранные белки, которые передают внеклеточные сигналы внутрь клетки. Рецепторы состоят из семи спиралей, окружающих мембрану. Активация рецептора агонистом способствует взаимодействию между рецептором и G-белком на внутренней поверхности мембраны. Это запускает обмен гуанозиндифосфата на гуанозинтрифосфат на α-субъединице G-белка и диссоциацию α-субъединицы от βγ-гетеродимера. Существует большое количество как активирующих, так и ингибирующих α-субъединиц, что приводит к разл. реакциям
Рецептор ГнРГ — рецептор, связанный с G-белком, который был выявлен в яичниках, яичках, гипоталамусе, предстательной железе, МЖ и плаценте (Yu, 2011). ГнРГ и его рецептор, вероятно, формируют аутокринную/паракринную регуляторную сеть в репродуктивных тканях, включая яичники и плаценту, в дополнение к классической нейроэндокринной гипоталамо-гипофизарной системе (Kim, 2007; Lee, 2010).
ЛГ и ХГЧ связываются с одним рецептором, сопряженным с G-белком, который носит название «рецептор ЛГ/ХГЧ». По сравнению с ЛГ ХГЧ обладает более выраженным сродством к рецептору и имеет более длительный период полураспада. ФСГ связывается с уникальным рецептором, сопряженным с G-белком, расположенным на мембране гранулезной клетки.
В яичнике рецепторы ЛГ/ХГЧ присутствуют на текаклетках, интерстициальных и лютеиновых клетках. В гранулезных клетках преантральных фолликулов молекулярная РНК рецептора ЛГ/ХГЧ практически не определяется. Однако в дифференцированных гранулезных клетках, обнаруженных во время созревания фолликула, определяются высокие уровни данного рецептора.
Рецепторы ЛГ/ХГЧ и ФСГ также обнаруживаются в эндометрии, миометрии и плаценте {Stilley, 2014; Ziecik, 2007). Функция рецептора в указанных экстраовариальных тканях изучена недостаточно.
б) Рецепторы стероидных гормонов:
1. Классификация и структура. Несмотря на структурное сходство, эстрогены, прогестагены, андрогены, глюко- и минералокортикоиды взаимодействуют с уникальными рецепторами, известными как гормональные ядерные рецепторы. Суперсемейство ядерных рецепторов состоит из трех групп. К ним относятся:
1) рецепторы, которые связывают стероидные лиганды;
2) рецепторы, которые имеют сродство к нестероидным лигандам, таким как гормон ЩЖ;
3) орфанные рецепторы.
Орфанный ядерный рецептор — рецептор, лиганд которого еще не открыт. Он считается конститутивно активным рецептором, т.е. проявляет базальную или внутреннюю активность.
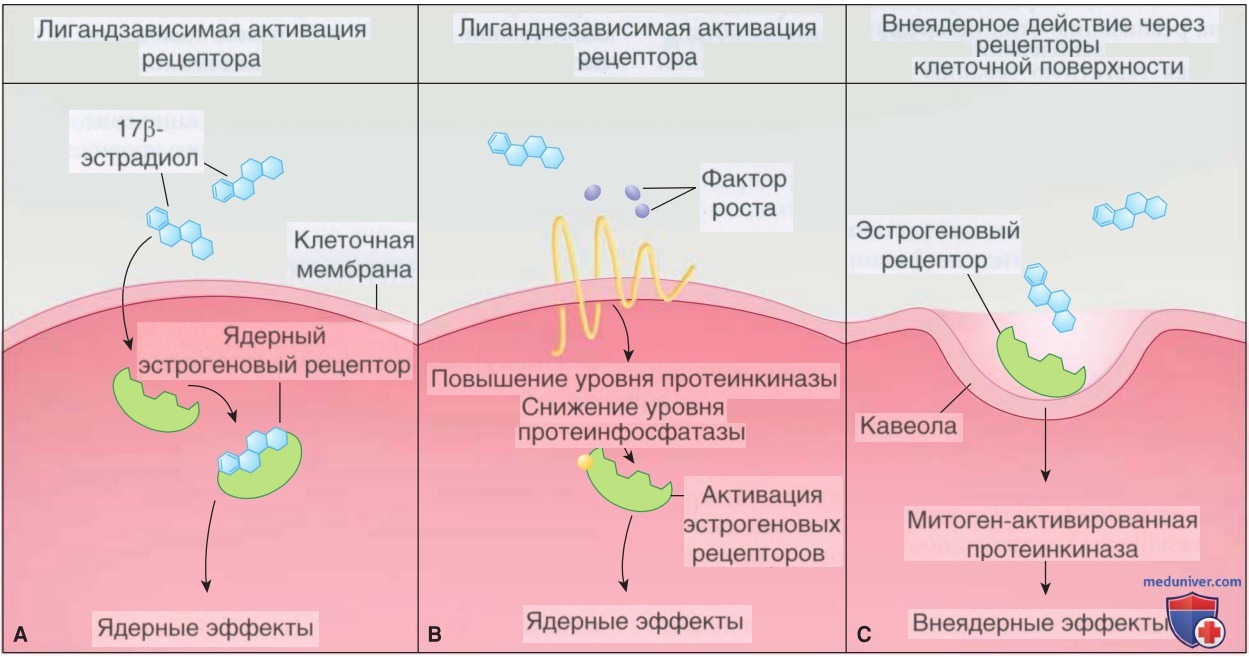
Свободные стероиды диффундируют в клетки и соединяются со специфическими рецепторами (рис. 2, А) (Gruber, 2002). Члены этого суперсемейства имеют модульную структуру разл. доменов (рис. 3) (O'Malley, 1999). Каждая группа отвечает за разл. виды деятельности, необходимые для полноценного функционирования рецептора.
Рисунок 2. Лигандзависимая и независимая активация эстрогеновых рецепторов: А — эстрогеновый рецептор м.б. активирован эстрогеном. Несвязанный гормон связывается со стероидными рецепторами, находящимися либо в цитоплазме, либо чаще всего в ядре клетки. Рецепторы, связанные с гормонами, затем связываются со специфическими промоторными последовательностями дезоксирибонуклеиновой кислоты. Это обычно приводит к транскрипции дезоксирибонуклеиновой кислоты и в конечном счете к синтезу специфического белка; В — эстрогеновый рецептор также м.б. активирован независимо от эстрогена. Факторы роста способствуют повышению активности протеинкиназ, которые фосфорилируют разл. участки на молекуле рецептора. Несвязанный, но активированный рецептор стимулирует транскрипционную активность; С — неядерные эстрогеновые сигнальные пути также являются триггерами. Эстрогеновые рецепторы расположены в инвагинациях клеточных мембран, называемых кавеолами. Связывание эстрогена с эстрогеновыми рецепторами проходит по митоген-активированному протеинкиназному пути и приводит к быстрому, внеядерному эффекту
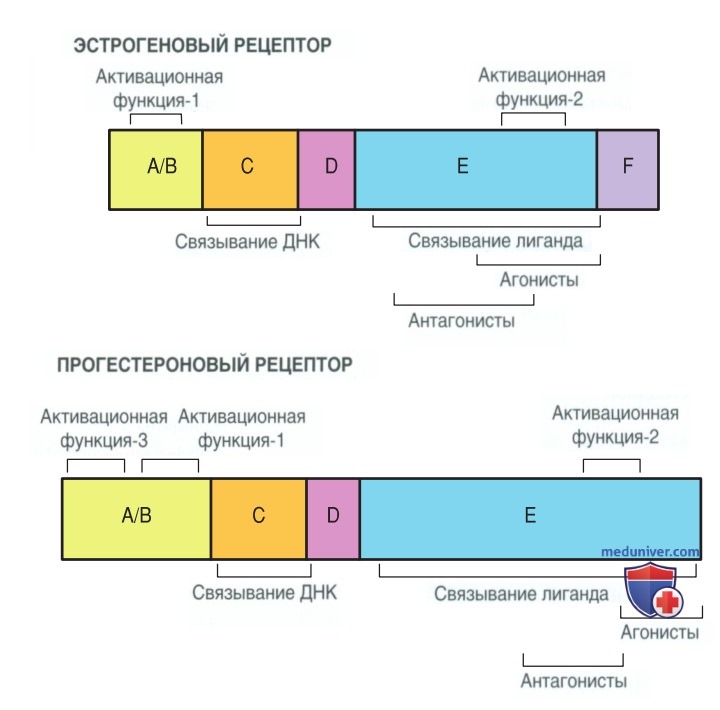
Рисунок 3. Функциональные домены внутри эстрогеновых и прогестероновых рецепторов, включая разл. сайты связывания лиганда и дезоксирибонуклеиновой кислоты
Установлено, что ядерные рецепторы содержат две области (участка), необходимые для активации гена, — активационную функцию-1 и активационную функцию-2. Активационная функция-1 находится в домене A/В и обычно не зависит от лиганда. Активационная функция-2 находится в лигандсвязывающем домене (Е) и часто является гормонозависимой. Консервативный участок связывания с ДНК (С) вставляется в спираль ДНК.
Впоследствии стероидные рецепторы усиливают или подавляют транскрипцию генов посредством взаимодействия со специфическими последовательностями ДНК, которые называются элементами гормонального ответа, в промоторной области генов-мишеней (Klinge, 2001).
2. Эстрогеновые, прогестероновые и андрогеновые рецепторы. Лиганднезависимые эстрогеновые рецепторы расположены в ядре. Прогестероновые рецепторы, андрогеновые рецепторы, а также рецепторы минерало- и глюкокортикоидов находятся в цитоплазме при отсутствии лиганда. Связывание лиганда с этими рецепторами позволяет осуществлять перемещение их в ядро.
Две изоформы эстрогеновых рецепторов α и β кодируются отдельными генами (Kuiper, 1997). Указанные рецепторы экспрессируются в разл. тканях и выполняют разные функции. Напр., эстрогеновые рецепторы α и β необходимы для нормального функционирования яичников. Отсутствие эстрогеновых рецепторов а у мышей приводит к ановуляции и накоплению кистозных фолликулов, в то время как недостаток эстрогеновых рецепторов β имеет нормальную гист. картину, несмотря на наличие ановуляторного цикла (Couse, 2000).
Прогестероновые рецепторы также существуют в нескольких изоформах. Прогестероновые рецепторы А и В кодируются одним геном и являются идентичными. Их структура различается только тем, что прогестероновый рецептор В содержит N-терминальный фрагмент из 164 аминокислот (Conneely, 2002). Подобно эстрогеновым рецепторам, изоформы прогестероновых рецепторов не являются взаимозаменяемыми.
Напр., прогестероновые рецепторы А необходимы для нормальной деятельности яичников и матки, но не влияют на функцию МЖ (Lydon, 1996). В отличие от эстрогеновых и прогестероновых рецепторов, была выявлена только одна форма андрогеновых рецепторов.
3. Негеномные действия стероидов. Подкласс стероидов, включая эстрогены и прогестины, может изменять функцию клеток через негеномный путь, т.е. независимо от классических ядерных рецепторов (см. рис. 2, С). Эти негеномные эффекты быстро развиваются и м.б. опосредованы через рецепторы на поверхности клеток, на примере с эстрогенами — трансмембранный сопряженный с G-белками рецептор эстрогена-30 (рецептор, сопряженный с G-белком) (GPR30) (Kowalik, 2013; Revelli, 1998).
Разрабатываемые ЛП специально нацелены на реализацию этих негеномных эффектов, обеспечивая таргетную терапию заболеваний репродуктивной системы (Tang, 2019).
в) Экспрессия рецепторов и десенсибилизация. Многие факторы изменяют клеточную реакцию на половые стероиды и пептидные факторы. Количество рецепторов внутри клетки или на клеточной мембране имеет решающее значение для достижения максимального гормонального ответа. Важно отметить, что количество рецепторов в клетке м.б. изменено путем транскрипции генов и деградации рецепторных белков.
Гормонально-индуцированная отрицательная обратная связь рецепторов называется отрицательной регуляцией, или десенсибилизацией. Десенсибилизация ограничивает продолжительность гормонального ответа, снижая чувствительность клетки к постоянному длительному действию гормона.
В репродуктивной системе десенсибилизация связана с рецепторами ГнРГ, что используется в клинической практике для создания гипоэстрогенного состояния. Агонисты ГнРГ, такие как лейпролида ацетат, первоначально стимулируют рецепторы гонадотрофов гипофиза, вызывая высвобождение ЛГ и ФСГ в большом количестве.
В течение нескольких часов агонисты подавляют гонадотропные рецепторы, тем самым десенсибилизируя их к дальнейшей стимуляции ГнРГ. Соответственно, снижение секреции гонадотропина приводит к уменьшению уровня эстрогена и прогестерона через 1-2 нед после первоначального введения агониста ГнРГ.