Менструальный цикл обычно длится 28±7 сут, продолжительность менструации составляет 4±2 сут с кровопотерей в среднем от 20 до 60 мл. Первым днем менструального цикла принято считать первый день вагинального кровотечения. Длительность менструального цикла может различаться среди женщин, а также может меняться в процессе репродуктивного периода жизни. Тем не менее цикл наиболее стабилен в возрасте от 20 до 40 лет.
По данным исследования, в котором приняли участие >2700 женщин, чаще отмечался нерегулярный менструальный цикл в течение 2 лет после менархе и за 3 года до менопаузы (Treloar, 1967). В частности, тенденция к сокращению длительности цикла с последующим его удлинением характерна для фазы менопаузального перехода. В недавнем исследовании было проанализировано >600 000 менструальных циклов, длительность цикла постепенно сокращалась на 0,19 сут в год у женщин в возрасте от 25 до 45 лет (Bull, 2019).
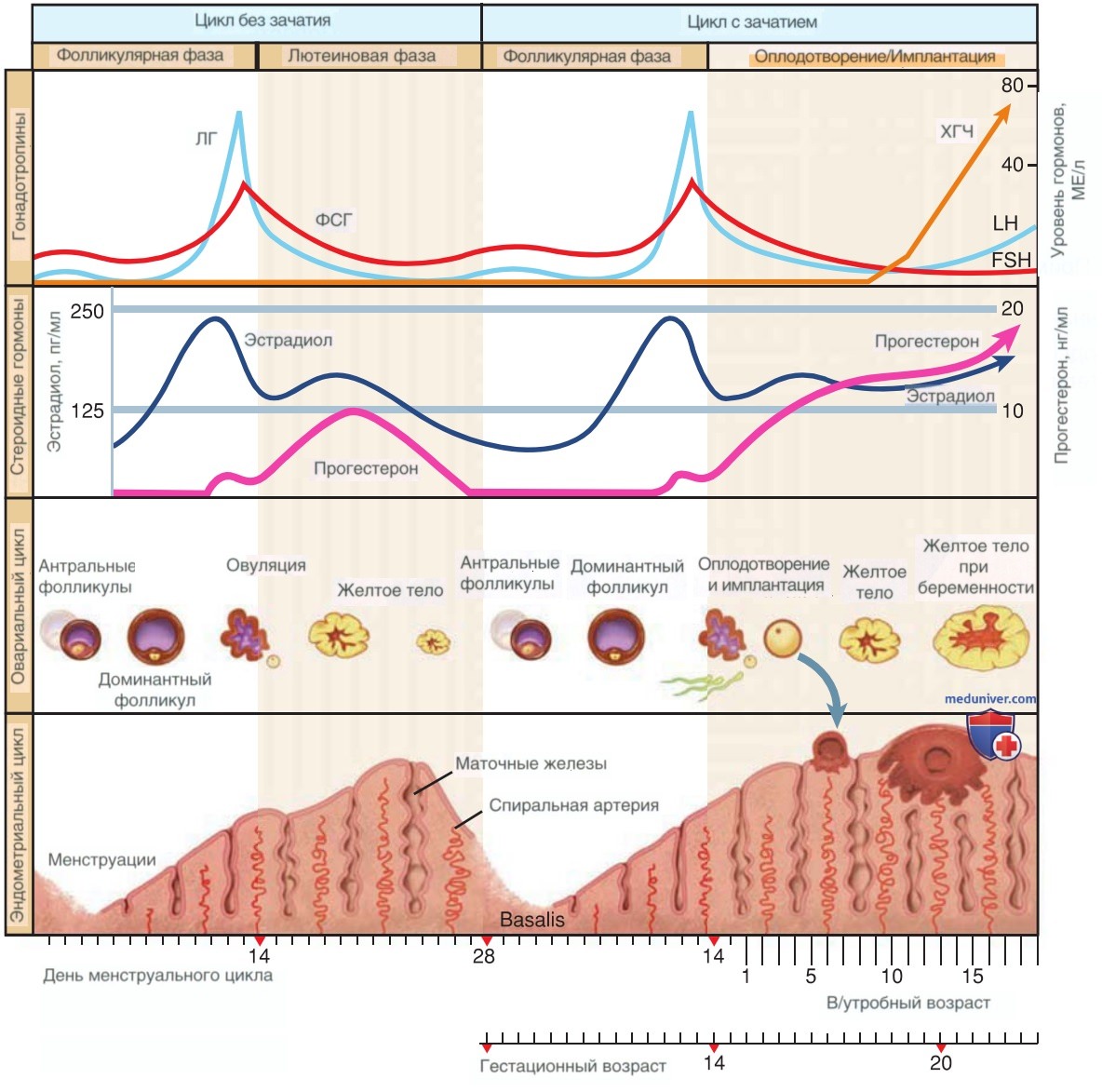
Яичник проходит следующие фазы менструального цикла: преовуляторную фолликулярную фазу и постовуляторную лютеиновую фазу, соответствующие фазы в эндометрии называются «пролиферативная» и «секреторная» (табл. 5). По результатам исследования средняя длительность фолликулярной фазы составила 16,9 сут, лютеиновой фазы — 12,4 сут (Bull, 2019). По мере появления новых данных может потребоваться переоценка классической длительности фазы цикла, но пока в этом нет необходимости (рис. 1).
Рисунок 1. Влияние гонадотропинов на овариальный и эндометриальный цикл. Продолжительность овариально-эндометриального цикла составляет 28 сут. При имплантации развивающаяся бластоциста начнет вырабатывать хорионический гонадотропин человека, сохраняя желтое тело, тем самым поддерживая выработку прогестерона
У большинства женщин лютеиновая фаза менструального цикла стабильна. Т.о., вариации в длительности цикла обычно являются результатом разной продолжительности фолликулярной фазы (Ferin, 1974).
а) Яичник:
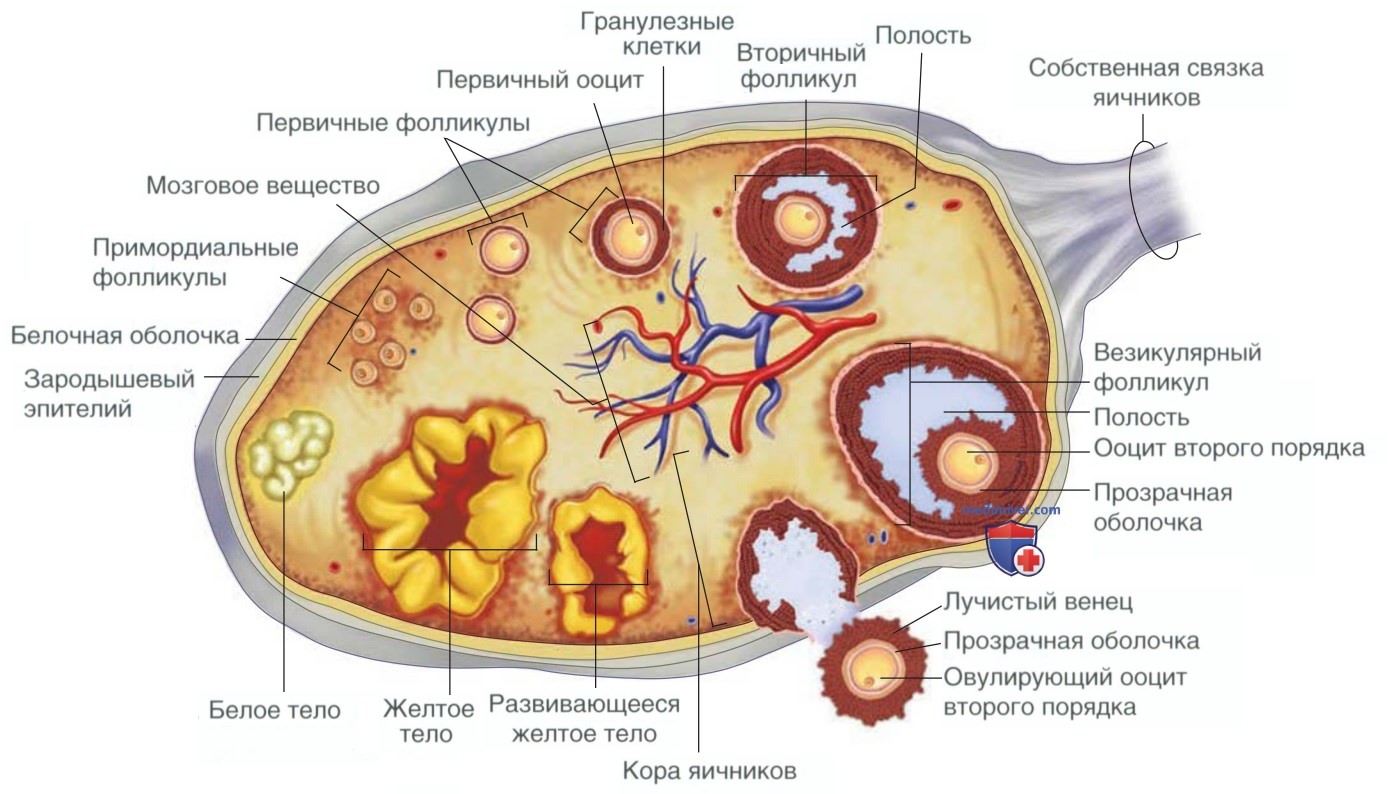
1. Морфология яичников. Яичник взрослой женщины овальной формы, длиной от 2 до 5 см, шириной от 1,5 до 3 см и толщиной от 0,5 до 1,5 см. В репродуктивные годы яичник весит от 5 до 10 г. Он состоит из трех частей. Внешний кортикальный слой содержит как зародышевый эпителий, так и фолликулы. Медуллярная область состоит из соединительной ткани, подобных миоидным сократимых клеток и интерстициальных клеток. Наконец, ворота яичника, в которые входят кровеносные, лимфатические сосуды и нервы (рис. 2).
Рисунок 2. Строение яичников и последовательные этапы развития фолликулов
Яичники выполняют две взаимосвязанные функции. Первая — выработка зрелых ооцитов. Вторая — выработка стероидных и пептидных гормонов, что создает среду, благоприятную для оплодотворения и последующей имплантации бластоцисты в эндометрий. В каждом цикле эндокринные функции яичника тесно связаны с появлением и исчезновением фолликулов и желтого тела.
2. Эмбриология яичников. В развитии яичника принимают участие три основных типа клеток. Во-первых, примордиальные клетки, которые развиваются из эндодермы желточного мешка, превращаются в первичные оогонии. Клетки целомического эпителия дают начало гранулезным клеткам, которые окружают ооциты. В-третьих, мезенхимальные клетки из полового гребня формируют строму яичника. Дополнительная информация о половой дифференциации содержится в отдельной статье на сайте (просим Вас пользоваться формой поиска по сайту выше).
Примордиальные клетки можно обнаружить в желточном мешке уже на 3-й неделе беременности (Gosden, 2013). Эти клетки мигрируют в половой гребень на 6-й неделе беременности и формируют первичный половой тяж. Яичники и семенники не отличаются по гист. критериям до 10-11 нед в/утробного периода.
Когда первичные клетки достигают гонады, они продолжают размножаться путем митотического деления. Начиная с 12 нед беременности часть оогоний входят в мейоз, превращаясь в первичные ооциты. Первичные ооциты в окружении одного слоя уплощенных гранулезных клеток образуют примордиальный фолликул.
3. Потеря яйцеклеток при старении. Все оогонии либо развиваются в первичные ооциты, либо подвергаются атрезии. Принято считать, что после рождения не происходит процесс созревания ооцитов. У мужчин, напротив, развитие сперматозоидов происходит непрерывно на протяжении всей взрослой жизни. Однако недавние исследования показывают, что стволовые клетки яичников могут генерировать зрелые яйцеклетки, что дает надежду на сохранение женской фертильности в будущем.
Однако в настоящее время эти заявления являются несколько спорными (Martin, 2019; Notarianni, 2011; Virant-Klun, 2015).
Максимальное количество оогоний (6-7 млн) достигается к 20-й неделе беременности (Baker, 1963). При рождении в яичнике присутствует от 1 до 2 млн оогоний. К началу периода полового созревания в яичнике остается <400 000 оогоний, из которых <500 в последующем овулируют. Т.о., в результате атрезии происходит потеря большинства половых клеток (Hsueh, 1996).
Фолликулярная атрезия представляет собой не пассивный некротический процесс, а активный процесс — апоптоз, находящийся под контролем гормональной регуляции. Апоптоз начинается в/утробно и продолжается в течение всего репродуктивного периода жизни.
4. Созревание яйцеклеток. Как уже упоминалось ранее, первичные оогонии, вступая в мейоз в/утробно, становятся первичными ооцитами. Прекращение развития ооцитов происходит в профазе I мейотического деления.
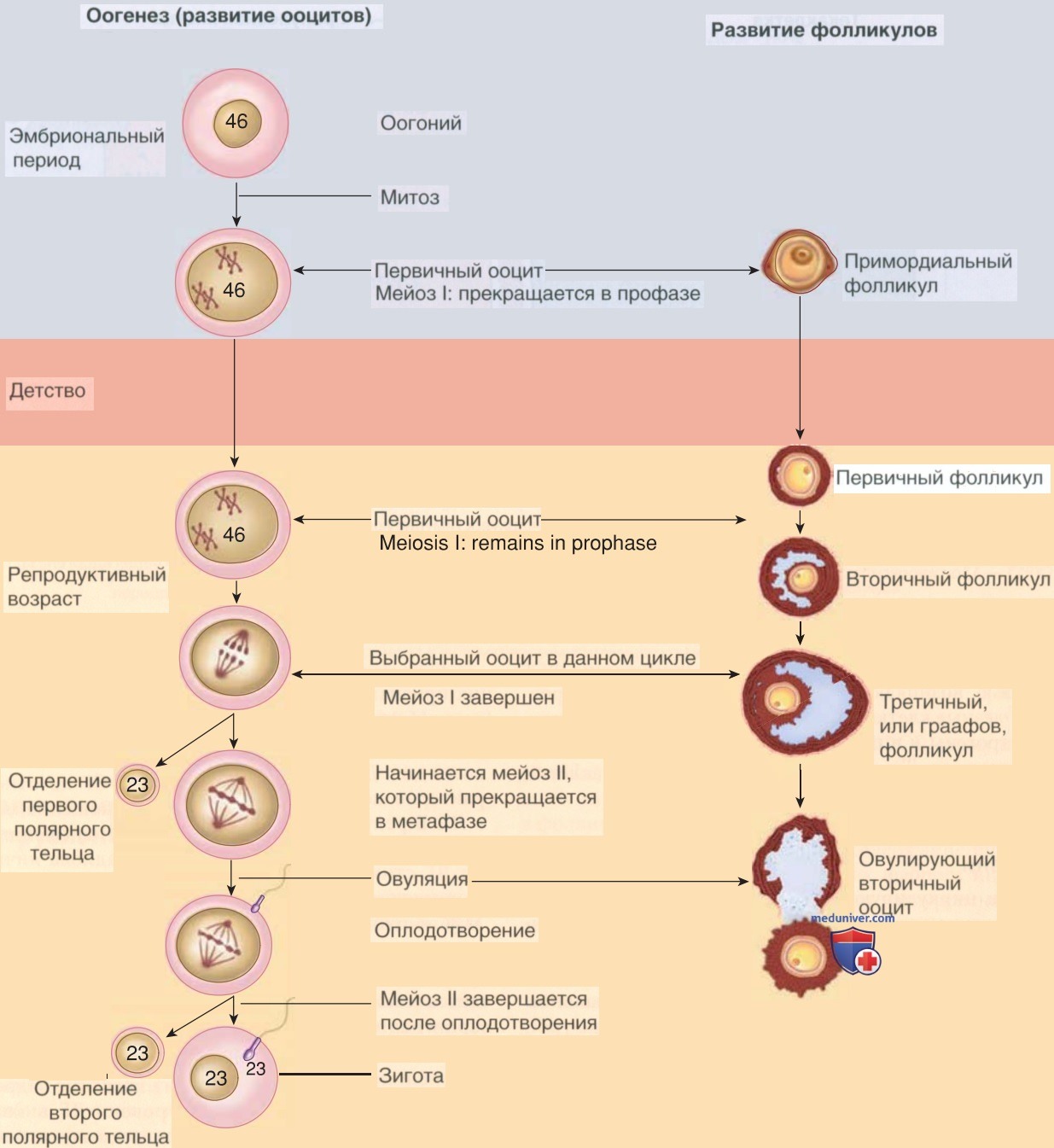
В когорте фолликулов возобновление мейоза происходит каждый месяц. Мейоз I завершается в ооците до овуляции в ответ на всплеск ЛГ. И снова процесс останавливается, на этот раз в метафазе II мейоза. Мейоз II завершается только в случае оплодотворения (рис. 3).
Рисунок 3. Этапы развития ооцита и созревания фолликулов. В эмбриональном периоде, достигая гонад, первичные половые клетки дифференцируются в оогонии. Количество оогоний продолжает увеличиваться путем митотического деления. Многие оогонии далее дифференцируются в первичные ооциты, с этого момента начинается мейоз. Однако процесс останавливается только после завершения профазы мейоза I. Первичный ооцит, окруженный эпителиальными клетками, называется примордиальным фолликулом. В детском возрасте первичные ооциты остаются в профазе I. Начиная с периода полового созревания и в течение репродуктивного периода несколько примордиальных фолликулов созревают каждый месяц в первичные фолликулы. Некоторые из них продолжают развиваться до вторичных фолликулов. Один или два вторичных фолликула развиваются в третичный, или граафов, фолликул. На этой стадии первое мейотическое деление завершается образованием гаплоидного вторичного ооцита и полярного тельца. Во время этого процесса цитоплазма сохраняется за счет вторичного ооцита. Следовательно, полярное тельце непропорционально мало. Вторичный ооцит останавливает мейоз на II профазе. Во время овуляции происходит высвобождение одного из вторичных ооцитов. II мейотическое деление завершается после оплодотворения яйцеклетки. При отсутствии оплодотворения яйцеклетка дегенерирует до завершения II мейотического деления
Помимо мейотического созревания, для нормального развития ооцитов необходимы изменения, происходящие в цитоплазме. Микротрубочки и актиновые нити позволяют перестроить клеточные органеллы, обеспечивая успешное отделение полярных телец и оплодотворение (Coticchio, 2015). Кумулюсные клетки, окружающие ооцит, модулируют созревание путем создания межклеточного контакта через щелевидное соединение и через секрецию паракринных факторов.
Понимание этих процессов и участвующих в них факторов способствует совершенствованию протоколов in vitro созревания ооцитов, что влияет на успешное сохранение фертильности и лечение бесплодия.
5. Стромальные клетки. Строма яичника содержит интерстициальные клетки, контрактильные клетки и клетки соединительной ткани. Клетки соединительной ткани способствуют поддержанию структуры яичника. Интерстициальные клетки, окружающие развивающийся фолликул, и гранулезные клетки дифференцируются в текаклетки. При гонадотропной стимуляции текаклетки увеличиваются в размере и накапливают запасы липидов, что характерно для стероидпродуцирующих клеток (Saxena, 1972).
Др. группа интерстициальных клеток в области ворот яичника носит название «хилусные клетки». Они очень похожи на клетки Лейдига, а гиперплазия или неопластические изменения хилусных клеток могут привести к избыточной секреции тестостерона и вирилизации. Роль этих клеток неизвестна, но их тесная связь с кровеносными сосудами и нейронами позволяет предположить, что они могут передавать системные сигналы в остальную часть яичника.
б) Выработка гормонов яичников. В норме яичник синтезирует и секретирует эстрогены, андрогены и прогестерон. Выработка гормонов находится под контролем гонадотропинов гипофиза, в частности ФСГ и ЛГ. Наиболее важными продуктами биосинтеза стероидов в яичниках являются прогестерон и эстрадиол. Однако яичник также выделяет большое количество эстрона, андростендиона, тестостерона и 17α-гидроксипрогестерона.
Половые стероидные гормоны играют важную роль в менструальном цикле, подготавливая матку к имплантации оплодотворенной яйцеклетки. При отсутствии имплантации происходят снижение уровня половых стероидов, дегенерация эндометрия и менструация.
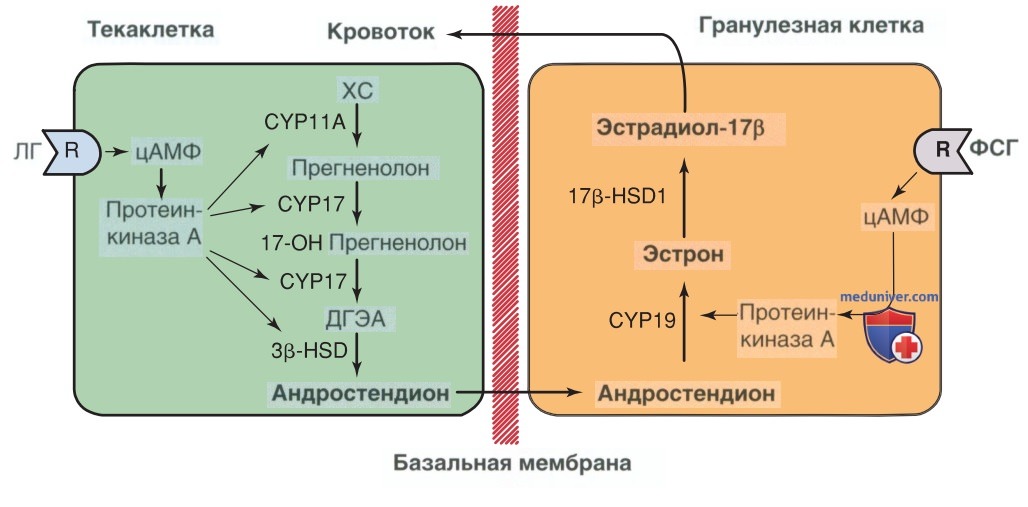
1. Двухклеточная теория. Для биосинтеза эстрогенов необходимо воздействие ЛГ и ФСГ на текаклетки и гранулезные клетки. Эта концепция известна как двухклеточная теория стероидогенеза яичников (рис. 4) (Peters, 1980). До поздней антральной стадии фолликулярного развития экспрессия ЛГ-рецепторов ограничена текаклетками, а экспрессия ФСГ-рецепторов — гранулезными клетками.
Рисунок 4. На рисунке схематично представлена теория двух клеток (или теория двух гонадотропинов) овариального стероидогенеза. Тека-клетки содержат большое количество рецепторов лютеинизирующего гормона. Связывание лютеинизирующего гормона с этими рецепторами приводит к активации циклического АМФК и синтезу андростендиона из холестерина. Андростендион диффундирует через базальную мембрану тека-клеток и проникает в гранулезные клетки яичника. Здесь при активации фолликулостимулирующего гормона, андростендион превращается с помощью фермента ароматазы в эстрон и эстрадиол. цАМФ — циклический аденозинмонофосфат; CYP11А — расщепляющий боковую цепь холестерина фермент; CYP17 — 17α-гидроксилаза; CYP19 — ароматаза; ДГЭА — дегидроэпиандростерон; 3β-HSD — Зр-гидроксистероиддегидрогеназа; 17β-HSD1 — 17β-гидроксистероиддегидрогеназа
Текаклетки выделяют все ферменты, необходимые для синтеза андростендиона, включая ген фермента CYP17, который катализирует 17-гидроксилирование. Это ограничивает скорость превращения прогестеронов в андрогены (Sasano, 1989). CYP17 отсутствует в гранулезных клетках, поэтому они не способны производить предшественников андрогенов, необходимых для синтеза эстрогенов, так что эту функцию выполняют текаклетки. В ответ на стимуляцию ЛГ текаклетки синтезируют андростендион и тестостерон.
Эти андрогены секретируются во внеклеточную жидкость и транспортируются через базальную мембрану к гранулезным клеткам, обеспечивая наличие предшественников для синтеза эстрогена. В отличие от тека-клеток, гранулезные клетки обладают высокой ароматазной активностью, возникающей в ответ на стимуляцию ФСГ. Т.о., гранулезные клетки эффективно преобразуют андрогены в эстрогены и в первую очередь в активный эстроген — эстрадиол. Т.о., стероидогенез яичников зависит от ЛГ и ФСГ, действующих независимо на текаклетки и гранулезные клетки соответственно.
2. Стероидогенез на протяжении всей жизни. Уровни циркулирующих гонадотропинов, ЛГ и ФСГ, заметно отличаются в разные возрастные периоды женщины. К 8-й неделе беременности яичник начинает вырабатывать эстрогены. Однако фактически во время в/утробного развития плода вырабатывается минимальное количество стероидов (Miller, 1988). Во II триместре уровень гонадотропинов в плазме повышается до уровня, характерного для менопаузы (Temeli, 1985).
В течение этого времени гипоталамо-гипофизарная ось плода продолжает созревать, становясь более чувствительной к высоким уровням циркулирующего эстрогена и прогестерона, секретируемых плацентой (Kaplan, 1976). До рождения в связи с высоким уровнем стероидов уровень гонадотропинов плода снижается.
После родов уровень гонадотропинов у новорожденного резко повышается в результате отделения плаценты и отсутствия ингибирования по типу обратной связи с плацентарными стероидами (Winter, 1976). Повышенный уровень гонадотропинов сохраняется в течение первых нескольких месяцев жизни, а затем снижается в раннем детском возрасте (Schmidt, 2000). Причины низкого уровня гонадотропина в этот период различны.
Гипоталамо-гипофизарная ось имеет повышенную чувствительность к отрицательной обратной связи, даже при низком уровне циркулирующих половых стероидов на этом этапе. Кроме того, ЦНС может поддерживать низкий уровень гонадотропинов. Низкие уровни ЛГ и ФСГ обнаруживаются даже у детей с дисгенезией гонад, у которых половые стероиды не реализуют механизм отрицательной обратной связи.
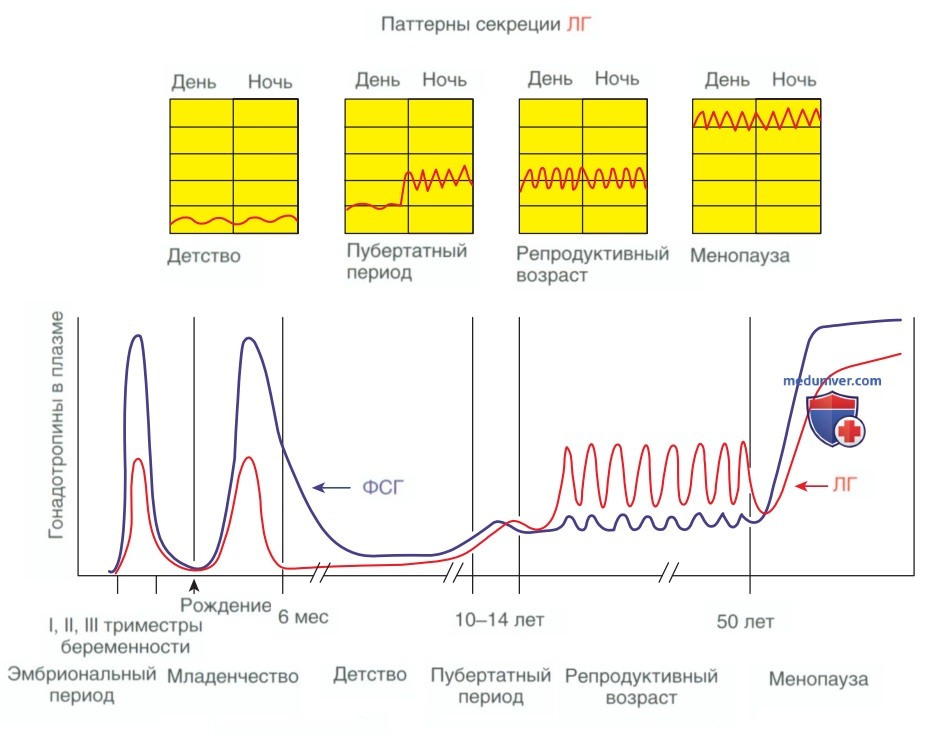
С наступлением полового созревания одним из ранних признаков является повышение секреции ЛГ во сне (рис. 5) (Faiman, 1976). Со временем отмечается увеличение секреции этого гонадотропина в течение всего дня. Для периода пременархе и постменопаузы характерно повышенное содержание ФСГ по отношению к ЛГ. В репродуктивный период соотношение меняется, уровень ЛГ превышает уровень ФСГ. Повышенные уровни ЛГ стимулируют выработку эстрадиола яичниками.
Рисунок 5. Разница в уровнях лютеинизирующего гормона и фолликулостимулирующего гормона в разл. возрастные периоды женщины
Повышение уровня эстрадиола приводит к увеличению МЖ в пубертатном периоде, которое называется «телархе», и созреванию внутренних и внешних половых органов. Формирование женского телосложения сочетается также и со скачком роста. Активация гипофизарно-надпочечниковой оси приводит к увеличению выработки андрогенов надпочечниками и пубертатным изменениям — появлению волос в лобковой и подмышечной зонах. Этот период носит название «адренархе» или «пубархе». Повышенный уровень ЛГ в конечном итоге приводит к овуляции и последующим менструациям.
Первая менструация называется «менархе». Этот процесс развития длится ~3-4 года и описывается далее в отдельной статье на сайте (просим Вас пользоваться формой поиска по сайту выше).
После менопаузы в яичнике остается лишь несколько фолликулов. В результате уровень эстрогенов и ингибинов в плазме крови заметно снижается после прекращения овуляции в яичнике. Из-за отсутствия отрицательной обратной связи уровни ЛГ и ФСГ резко повышаются. Высокий уровень ЛГ может стимулировать выработку С19-стероидных гормонов (главным образом андростендиона) в стромальных клетках яичников. Периферические ткани могут преобразовывать синтезируемый яичниками андростендион и андрогены надпочечников в эстрон — основной эстроген в сыворотке крови у женщин в постменопаузальном периоде.
Андростендион превращается в эстрон в основном в жировой ткани. Синтез эстрона из андростендиона напрямую зависит от массы тела. При одинаковой массе тела количество синтезируемого эстрона больше у женщин в постменопаузе, чем у женщин в пременопаузе. Однако низкий уровень циркулирующего эстрогена обычно не обеспечивает защиты от потери плотности костной ткани.
в) Гонадные пептиды и менструальный цикл. Ингибин, активин и фоллистатин являются гонадными пептидами, которые регулируют гонадотропную активность (de Kretser, 2002). Как следует из их названий, ингибин подавляет, а активин стимулирует гонадотропную функцию. Фоллистатин подавляет экспрессию гена FSHβ, вероятно, связываясь с активином и тем самым предотвращая его взаимодействие с рецептором (Xia, 2009).
Ингибин и активин — два тесно связанных пептида. Ингибин состоит из α-субъединицы (не связанной с α-субъединицей гликопротеина ЛГ и ФСГ), связанной дисульфидным мостиком с одной из двух высокогомологичных beta;-субъединиц с образованием ингибина А (αbeta;А) или ингибина В (αbeta;В). Активин состоит из гомодимеров (beta;Abeta;A, beta;Bbeta;B) или гетеродимеров (beta;Аbeta;В) тех же beta;-субъединиц, что и ингибин (Bilezikjian, 2012). Напротив, фоллистатин структурно отличается от ингибина и активина.
Хотя первоначально эти пептиды были выделены из фолликулярной жидкости, они вырабатываются в гипофизе, яичниках, семенниках и плаценте, а также в ГМ, надпочечниках, печени, почках и костном мозге, выполняя разл. тканеспецифические функции (Muttukrishna, 2004). Активин и фоллистатин, скорее всего, действуют как аутокринные/паракринные факторы в тканях, в которых они вырабатываются, включая яичник.
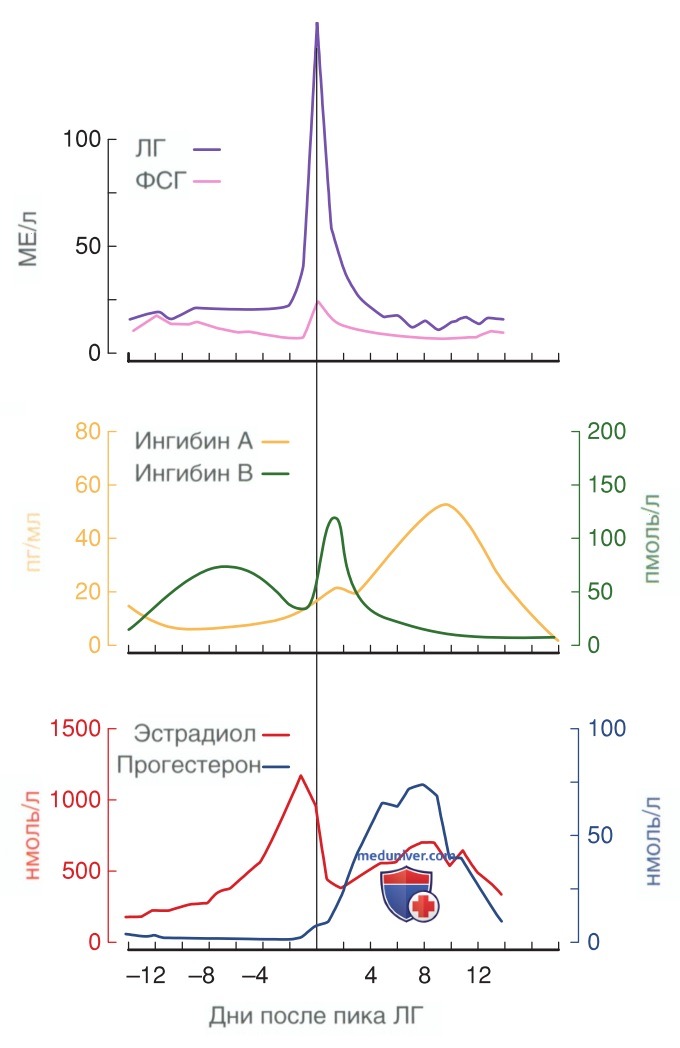
Напротив, ингибины, продуцируемые яичниками, циркулируют в значительных концентрациях и, вероятно, имеют решающее значение в отрицательной обратной связи генной экспрессии гонадотропина. В частности, во время ранней фолликулярной фазы ФСГ стимулирует секрецию ингибина В гранулезными клетками (рис. 6) (Buckler, 1989). Однако повышение уровня циркулирующего ингибина В тормозит последующую секрецию ФСГ в фолликулярной фазе.
Рисунок 6. Динамика уровней гонадотропинов, ингибина и половых стероидов на протяжении нормального менструального цикла. Верхний график показывает пик уровня лютеинизирующего гормона (фиолетовая линия) и фолликулостимулирующего гормона (розовая линия). Средний график показывает изменение уровней ингибина А и ингибина В. Обратите внимание, что уровень ингибина В (зеленая линия) достигает пика по времени рядом с всплеском уровня лютеинизирующего гормона в середине цикла, тогда как максимальное повышение уровня ингибина А (оранжевая линия) происходит через несколько суток после этого пика. На нижнем графике отмечено повышение уровня эстрадиола (красная линия) до всплеска уровня лютеинизирующего гормона и в середине лютеиновой фазы. Уровень прогестерона (синяя линия) достигает пика в середине лютеиновой фазы
Во время лютеиновой фазы ЛГ регулирует выработку ингибина, и содержание ингибина А повышается (McLachlan, 1989). Пик выброса ингибина В наблюдается при всплеске ЛГ, тогда как уровень ингибина А достигает максимальных значений через несколько дней, в середине лютеиновой фазы. Уровни ингибина снижаются при завершении лютеиновой фазы и остаются низкими во время перехода в фолликулярную фазу и в ранней фолликулярной фазе. Обратная зависимость между уровнем ингибина и ФСГ в крови согласуется с ролью ингибина в регуляции секреции ФСГ по принципу отрицательной обратной связи.
Инсулиноподобные факторы роста также влияют на регуляцию функции яичников. Только инсулиноподобный фактор роста-2 участвует в развитии первичных фолликулов. Оба инсулиноподобных фактора роста-1 и 2 стимулируют рост вторичных фолликулов. Гонадотропины стимулируют выработку инсулиноподобного фактора роста-2 в текаклетках, клетках гранулезы и лютеинизированных клетках гранулезы.
Рецепторы инсулиноподобного фактора роста экспрессируются в текаклетках и клетках гранулезы, поддерживая аутокринное/паракринное действие в фолликуле. ФСГ также влияет на экспрессию инсулиноподобных факторов роста. Эта комплексная система помогает регулировать интрафолликулярную активность (Silva, 2009).
г) Развитие фолликулов:
1. Стадии развития фолликула. Фолликулярное развитие начинается с примордиальных фолликулов, которые были сформированы в течение внутриутробной жизни плода (см. рис. 3). Эти фолликулы состоят из ооцита, развитие которого приостановлено на стадии первого мейотического деления, окруженного одним слоем уплощенных гранулезных клеток. Фолликулы отделены от стромы тонкой базальной мембраной. Преовуляторные фолликулы являются аваскулярными, в связи с чем их рост полностью зависит от диффузии питательных веществ через щелевидные контакты, посредством которых также происходит выход продуктов метаболизма.
Кроме того, путем диффузии также осуществляется транспорт предшественников стероидных гормонов из слоя текаклеток в гранулезный слой.
На стадии первичного фолликула гранулезные клетки становятся кубовидными, их количество увеличивается, образуя псевдостратифицированный (зернистый) слой. Межклеточные щелевидные контакты находятся между соседними гранулезными клетками, а также между гранулезными клетками и развивающимся ооцитом (Albertini, 1974). Щелевидные контакты позволяют клеткам при отсутствии рецепторов к гонадотропинам получать сигналы от клеток, экспрессирующих эти рецепторы, что способствует передачи гормональных эффектов фолликулу.
На этой стадии яйцеклетка выделяет в-ва, образующие внеклеточную оболочку, известную как zona pellucida, которая содержит по крайней мере три белка: ZP1, ZP2 и ZP3. Рецепторы на головке акросомы сперматозоида взаимодействуют с ZP3. При взаимодействии выпускается содержимое акросомы, происходят пенетрация zona pellucida и оплодотворение. Ферменты, высвобождаемые из акросомы, провоцируют изменения в ZP2, приводящие к затвердению оболочки, что предотвращает оплодотворение яйцеклетки более чем одним сперматозоидом (Gupta, 2015).
Развитие вторичного, или преантрального, фолликула включает конечный рост ооцита и дальнейшее увеличение числа гранулезных клеток. Слой стромальных клеток, окружающих клетки гранулезы, можно разделить на theca interna и theca externa (Eppig, 1979).
Третичные фолликулы, также называемые антральными фолликулами, образуются в результате продолжения развития ооцита. В них фолликулярная жидкость собирается внутри гранулезных клеток, в конечном счете образуя заполненную жидкостью полость. Гранулезные клетки в антральном фолликуле гистологически и функционально делятся на две группы. Гранулезные клетки, окружающие ооцит, образуют яйценосный бугорок, тогда как гранулезные клетки, окружающие полость, носят название «пристеночные гранулезные клетки».
Антральная жидкость состоит из плазматического фильтрата и факторов, секретируемых клетками гранулезы. Эти факторы включают эстроген и факторы роста, они присутствуют в значительно более высоких концентрациях в фолликулярной жидкости, чем в крови, и, вероятно, имеют решающее значение для успешного созревания фолликулов (Asimakopoulos, 2006; Silva, 2009). Дальнейшее накопление антральной жидкости приводит к быстрому росту фолликула и развитию преовуляторного фолликула, также называемого «граафов фолликул» (Hennet, 2012).
Во время этого процесса на ранней стадии развития (вплоть до вторичного фолликула) гонадотропная стимуляция не требуется, и поэтому фолликулы считаются «гонадотропиннезависимыми». Для окончательного созревания фолликулов требуется достаточное количество циркулирующих гонадотропинов (ЛГ и ФСГ), и поэтому на этой стадии они считаются «гонадотропинзависимыми» (Butt, 1970). Две эти стадии накладываются друг на друга, хотя ранее считалось, что происходят в два отдельных этапа.
2. Окно отбора (селекция фолликула). Фолликулогенез — многоступенчатый процесс, который продолжается не менее 3 мес и завершается овуляцией фолликула. Каждый месяц в когорте фолликулов начинается фаза полусинхронного роста. Размер когорты, вероятно, зависит от количества неактивных примордиальных фолликулов в яичниках и у молодых женщин состоит от 3 до 11 фолликулов в каждом яичнике (Hodgen, 1982; Pache, 1990). Важно отметить, что овуляторный фолликул рекрутируется из когорты фолликулов, начавших свое развитие за 2-3 предшествующих цикла, тогда как большинство фолликулов подвергается атрезии.
Во время поздней лютеиновой фазы повышение уровня ФСГ способствует рекрутированию единственного доминирующего фолликула, который в конечном счете овулирует (Schipper, 1998). Как было описано ранее, тека-клетки вырабатывают андрогены, а гранулезные клетки — эстрадиол. Уровень эстрадиола повышается с ростом фолликула, что способствует усилению воздействия ФСГ на гранулезные клетки и создает положительную прямую связь с фолликулами, которые продуцируют эстрадиол.
Считается, что вырабатываемые яичником инсулиноподобные факторы роста, взаимодействуя с ФСГ, оказывают влияние на выбор доминирующего фолликула (Son, 2011). Кроме того, в/яичниковое повышение уровня VEGF потенцирует эффекты ФСГ путем усиления васкуляризации доминирующего фолликула.
Выработка доминирующим фолликулом эстрадиола, а также ингибина В гранулезными клетками способствует подавлению уровня ФСГ в фолликулярной фазе, что может препятствовать достижению преовуляторного статуса др. фолликулов в течение цикла.
3. Эстроген-доминирующее микроокружение. Для продолжения созревания фолликула требуется успешный переход от «андроген-доминирующего» микроокружения к «эстроген-доминирующему». При низких концентрациях андрогены стимулируют ароматизацию и способствуют выработке эстрогена. Однако в/фолликулярный уровень андрогенов будет повышаться, если выработка андрогенов слоем текаклеток происходит быстрее ароматизации в клетках гранулезы. При более высоких концентрациях андрогены превращаются в более активные 5α-андрогены, такие как ДГТ.
Эти андрогены ингибируют активность ароматазы, не м.б. ароматизированы до эстрогенов и ингибируют ФСГ-индукцию экспрессии рецепторов ЛГ на гранулезных клетках (Gervdsio, 2014).
Согласно этой модели, фолликулы, которые не имеют надлежащего рецептора ФСГ и достаточного количества гранулезных клеток, останутся в основном андрогенными и станут атретическими. В фолликулярной жидкости атретических фолликулов обнаружено повышенное соотношение андрогенов и эстрогенов, и, по данным нескольких исследований, высокий уровень эстрогенов предотвращает апоптоз.
Инсулиноподобный фактор роста-1 также подавляет активность апоптоза и продуцируется клетками гранулезы. Этот эффект ИФР-1 ослабляется определенными ИФР-связывающими белками, которые присутствуют в фолликулярной жидкости атретических фолликулов. Таким образом, предотвращение атрезии ФСГ м.б. частично обусловлено его способностью стимулировать синтез инсулиноподобного фактора роста-1 и подавлять синтез связывающих инсулиноподобные факторы роста белков.
д) Фазы менструального цикла:
1. Фолликулярная фаза. В поздней лютеиновой фазе уровни эстрадиола, прогестерона и ингибина резко снижаются, тогда как уровень циркулирующего ФСГ повышается (Hodgen, 1982). Как было упомянуто ранее, высокий уровень ФСГ стимулирует рекрутирование когорты фолликулов, в результате которого один из них доходит до предовуляторной стадии. Согласно данным УЗИ, овуляция происходит в яичниках случайным образом (Baird, 1987).
У женщин со сниженной функцией яичников уровень ФСГ в ранней фолликулярной фазе повышен по сравнению с молодыми женщинами с нормальной овариальной функцией, предположительно из-за снижения выработки ингибина яичниками в предшествующей лютеиновой фазе, поэтому традиционно уровень ФСГ и эстрадиола измеряют на 3-й день цикла. Ускоренное повышение уровня ФСГ в сыворотке крови приводит к более активному рекрутированию фолликулов. Это объясняет сокращение фолликулярной фазы у женщин позднего репродуктивного возраста и повышение вероятности рождения близнецов.
В середине фолликулярной фазы фолликулы вырабатывают большее количество эстрадиола и ингибина. Это приводит к снижению уровня ФСГ по типу отрицательной обратной связи. Считается, что снижение уровня ФСГ способствует селекции фолликула, предназначенного для овуляции. Такой фолликул называется доминирующим. Исходя из этой теории, оставшиеся фолликулы, которые подвергнутся атрезии, экспрессируют меньше рецепторов ФСГ и не способны реагировать на снижение уровня ФСГ.
Как отмечалось ранее, уменьшение экспрессии VEGF также играет определенную роль за счет снижения васкуляризации и транспорта циркулирующих факторов к этим фолликулам (Ravindranath, 1992).
В течение большей части фолликулярного развития гранулезные клетки реагируют на ФСГ путем увеличения их количества, экспрессии ароматазы и под действием эстрадиола — экспрессии рецепторов ЛГ на их поверхности. Под действием экспрессирующегося рецепторами ЛГ в поздней фолликулярной фазе гранулезные клетки начинают вырабатывать небольшое количество прогестерона. Прогестерон уменьшает пролиферацию гранулезных клеток и тем самым замедляет рост фолликулов (Chaffkin, 1992).
2. Овуляция. К концу фолликулярной фазы уровень эстрадиола резко возрастает, что способствует переходу отрицательной в положительную обратную связь как в гипоталамусе, так и в передней доле гипофиза. Этот процесс не до конца изучен и, возможно, связан с изменениями в нейронах кисспептина. Такое изменение обратной связи вызывает пик уровня ЛГ. При этом для пика ЛГ необходимо, чтобы концентрация эстрадиола достигала концентрации 200 пг/мл в течение 50 ч (Young, 1976).
Небольшое предовуляторное повышение концентрации прогестерона вызывает всплеск уровня ФСГ, который происходит вместе с пиком ЛГ (McNatty, 1979). Прогестерон также может потенцировать способность эстрадиола к стимуляции пика ЛГ. Это объясняет внезапное начало овуляции у женщин с ановуляторной аменореей при приеме прогестерона для стимуляции менструации.
Пик ЛГ быстро воздействует как на гранулезные, так и на текаклетки преовуляторного фолликула, прекращая экспрессию генов, участвующих в фолликулярной экспрессии, и активируя гены, необходимые для овуляции и лютеинизации. Кроме того, всплеск ЛГ инициирует возвращение яйцеклетки в мейоз, расширение яйценосного бугорка, синтез Pg и лютеинизацию гранулезных клеток. Средняя продолжительность всплеска ЛГ составляет 48 ч, тогда как овуляция происходит через 36-40 ч после начала всплеска ЛГ (Hoff, 1983; Lemarchand-Beraud, 1982).
Повышение уровня ЛГ резко прекращается, вероятно, после резкого увеличения секреции стероидов и ингибинов желтым телом. В качестве альтернативы также может потребоваться выработка яичником либо гипоталамусом фактора (GnSIFlAF), усиливающего или ослабляющего высвобождение гормона гонадотропина. Однако характеристики этого фактора неизвестны (Vega, 2015).
Гранулезные клетки, окружающие ооцит, в отличие от муральных гранулезных клеток, не экспрессируют ЛГ-рецепторы и не синтезируют прогестерон. Между этими гранулезными клетками яйценосного бугорка и яйцеклеткой развиваются плотные щелевидные соединения. Шероховатая поверхность и довольно большой размер яйценосного бугорка способствуют «захвату» яйцеклетки трубчатыми фимбриями.
Недавно были открыты эпидермальные факторы роста — амфирегулин, эпирегулин и бетацеллюлин, — которые под влиянием ЛГ могут способствовать морфологическим и биохимическим изменениям в яичнике (Hsieh, 2009), приводящим к овуляции.
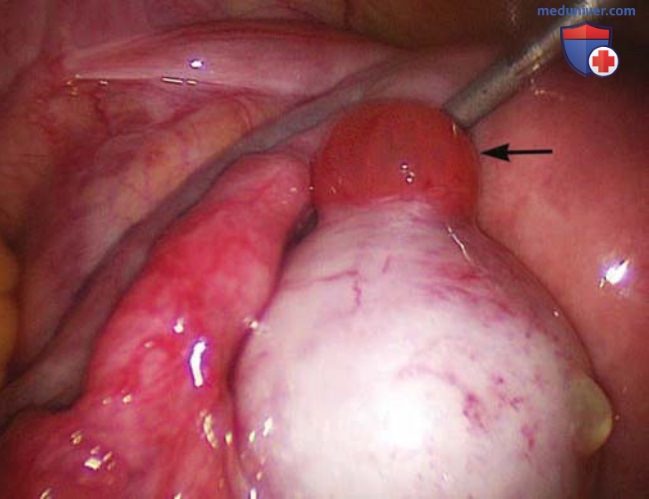
Согласно данным УЗИ, экструзия ооцита длится всего несколько минут (рис. 7) (Knobil, 1994). Этот механизм недостаточно изучен, однако не связан с повышением давления в фолликуле (Espey, 1974). Под действием протеолитических ферментов в фолликуле, включая плазмин и коллагеназу, происходит истончение фолликулярной стенки (Beers, 1975). Преовуляторный пик ЛГ стимулирует экспрессию тканевого активатора плазминогена гранулезными и текаклетками, а также экспрессию ингибитора плазминогена, приводя к выраженному повышению активности плазминогена (Piquette, 1993).
Рисунок 7. Во время лапароскопии на поверхности яичника до овуляции визуализируется стигма
Pg также достигают пиковой концентрации в фолликулярной жидкости во время преовуляторного пика ЛГ (Lumsden, 1986). Pg могут стимулировать сокращение гладкой мускулатуры яичника и тем самым способствовать овуляции (Yoshimura, 1987). Женщинам, проходящим лечение бесплодия, не рекомендуется применять ингибиторы простагландинсинтетазы в преовуляторный период во избежание синдрома лютеинизации неовулировавшего фолликула (Smith, 1996). Неизвестно, является ли лютеинизация неовулировавшего фолликула патологическим синдромом или имеет спорадическое течение (Kerin, 1983).
3. Лютеиновая фаза. После овуляции оставшиеся клетки фолликула дифференцируются в желтое тело (Corner, 1956). Этот процесс, требующий стимуляции ЛГ, включает в себя как морфологические, так и функциональные изменения и известен как «лютеинизация». Вследствие пролиферации и гипертрофии гранулезных и текаклеток образуются гранулезнолютеиновые клетки и более мелкие текалютеиновые клетки соответственно (Patton, 1991).
Во время формирования желтого тела происходит дегенерация базальной мембраны, отделяющей гранулезные клетки от текаклеток, а также через 2 сут после овуляции — васкуляризация ранее аваскулярных гранулезных клеток. Капилляры достигают центра желтого тела к 4-му дню. Вследствие усиления перфузии лютеиновые клетки получают доступ к циркулирующему ЛПНП, который содержит необходимый для биосинтеза стероидов ХС. Усиление кровоснабжения также может иметь клинические последствия. Наиболее частой жалобой при обращении в ОНМП является боль от геморрагической кисты желтого тела.
Стероидогенез в желтом теле зависит от уровня ЛГ в сыворотке крови, ЛГ-рецепторов на лютеиновых клетках и достаточного количества лютеиновых клеток. Так, крайне важно, чтобы экспрессия ЛГ-рецепторов на клетках гранулезы была надлежащим образом индуцирована во время предыдущей фолликулярной фазы. Снижение концентрации ЛГ в сыворотке крови связано с укороченной лютеиновой фазой. На лютеиновую функцию также влияют уровни гонадотропинов предыдущей фолликулярной фазы. Снижение секреции ЛГ или ФСГ связано с нарушением лютеиновой функции. Недостаток ФСГ, вероятно, приводит к уменьшению количества гранулезных клеток.
Кроме того, лютеиновые клетки при таком недостаточно эффективном цикле имеют меньше ФСГ-индуцированных рецепторов ЛГ и, следовательно, будут менее чувствительны к стимуляции ЛГ.
Максимальная концентрация прогестерона наблюдается в лютеиновую фазу цикла, тогда как в фолликулярной фазе происходит активный синтез эстрадиола. Повышенная васкуляризация, гипертрофия клеток и увеличение количества в/клеточных органелл приводят к преобразованию желтого тела в наиболее активную стероидогенную ткань в организме. Секреция максимальных уровней прогестерона наблюдается в середине лютеиновой фазы и составляет 40 мг/сут. Можно с уверенностью предположить, что овуляция произошла, если уровень прогестерона >3 нг/мл на 21-й день цикла.
Хотя прогестерон является главным стероидом, вырабатываемым яичниками во время лютеиновой фазы, эстрадиол также вырабатывается в значительном количестве. Уровень эстрадиола быстро падает сразу после всплеска ЛГ, что объясняет появление мажущих кровянистых выделений в середине цикла у некоторых женщин. Хотя причина неизвестна, снижение уровня эстрадиола м.б. результатом прямого ингибирования роста гранулезных клеток вследствие повышения уровня прогестерона (Hoff, 1983). Вслед за снижением уровень эстрадиола неуклонно повышается и достигает максимума в середине лютеиновой фазы.
Желтое тело также синтезирует большое количество ингибина А. Одновременно происходит падение уровня циркулирующего ФСГ в лютеиновой фазе. Если уровень ингибина А снижается в конце лютеиновой фазы, уровень ФСГ снова повышается, и начинается отбор когорты ооцитов для следующего менструального цикла.
При отсутствии наступления беременности происходит разрушение желтого тела — лютеолиз. Этот механизм плохо изучен, но предполагается, что лютеиновая регрессия является строго регулируемым процессом. После лютеолиза кровоснабжение желтого тела уменьшается, секреция прогестерона и эстрадиола резко падает, а лютеиновые клетки подвергаются апоптозу и становятся фиброзными, приводя к формированию белого тела.
При наступлении беременности ХГЧ, вырабатываемый на ранних сроках беременности, «защищает» желтое тело от атрезии, связываясь с ЛГ-рецептором на лютеиновых клетках и активируя его. Стимуляция стероидогенеза желтого тела гормоном ХГЧ помогает поддерживать стабильность эндометрия до тех пор, пока выработка половых стероидов плацентой не станет достаточно стабильной для выполнения этой функции в конце I триместра.
По этой причине при хирургическом удалении желтого тела во время беременности необходимо вводить прогестерон до ~10 нед беременности, как описано в отдельной статье на сайте (просим Вас пользоваться формой поиска по сайту выше).