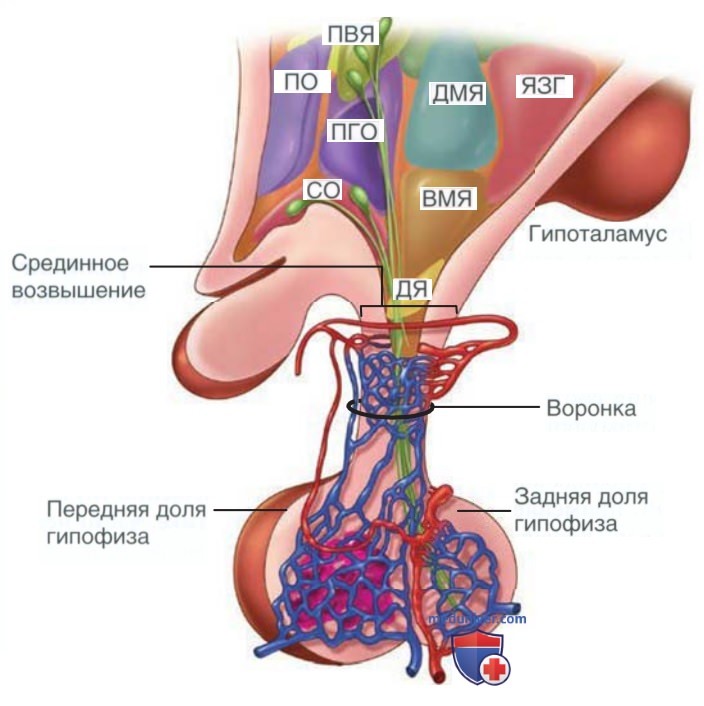
а) Строение. Гипоталамус состоит из ядер, расположенных в основании ГМ, чуть выше хиазмы зрительных нервов. Нейроны гипоталамуса образуют синаптические связи с нейронами ЦНС. Часть гипоталамических нейронов в дугообразном, вентромедиальном и паравентрикулярном ядрах проецируются на срединное возвышение. В срединном возвышении из верхних гипофизарных артерий формируется плотная капиллярная сеть (рис. 1).
Рисунок 1. Сагиттальный срез гипоталамуса и гипофиза со структурами ростральной зоны слева и каудальной — справа. Анатомически и функционально гипоталамус связан с передней долей гипофиза портальной системой кровоснабжения. Задняя доля гипофиза содержит нервные окончания аксонов, принадлежащие нейронам супраоптического (СО) и паравентрикулярного ядер (ПВЯ) гипоталамуса: ПГО — передняя гипоталамическая область; ДЯ — дугообразное ядро; ДМЯ — дорсомедиальное ядро; ЯЗГ — ядра заднего гипоталамуса; ПО — преоптическая область; ВМЯ — вентромедиальное ядро
Капилляры впадают в портальные сосуды, которые пересекают ножку гипофиза, образуя капиллярную сеть в передней доле гипофиза (аденогипофиз). Портальная система кровообращения гипофиза в основном направлена от гипоталамуса к гипофизу. Однако также существует обратный кровоток. Это создает сверхкороткую петлю обратной связи между гипофизом и гипоталамическими нейронами. Гипоталамус является основным пространством интеграции информации, поступающей из окружающей среды, НС и многих систем органов.
Передняя доля гипофиза состоит из эндокринных клеток и развивается в результате выпячивания стенки ротовой бухты — кармана Ратке. Задняя доля гипофиза (нейрогипофиз) представляет собой нервную ткань и состоит из нервных окончаний аксонов, принадлежащих крупноклеточным нейронам супраоптического и паравентрикулярного ядер гипоталамуса.
б) Нейроэндокринология гипоталамуса. Список известных нейротрансмиттеров увеличивается по мере того, как растет понимание их анатом. распределения, способа регуляции и механизма действия. Нейротрансмиттеры можно разделить на следующие группы: 1) биогенные амины (дофамин, адреналин, норадреналин, серотонин, гистамин); 2) нейропептиды; 3) ацетилхолин; 4) возбуждающие аминонейротрансмиттеры (глутамат, глицин, аспарагиновая кислота); 5) тормозной нейромедиатор ГАМК; 6) газовые трансмиттеры (оксид азота, монооксид углерода) и 7) др. факторы (цитокины, факторы роста).
Клинически важные нейропептиды, регулирующие репродуктивную систему, включают эндогенные опиаты, кисспептин, галанин, нейрокинин В, нейропептид Y и пептид, активирующий аденилатциклазу гипофиза. Этот список быстро растет по мере продолжения исследований и не является конечным.
1. Эндогенные опиаты. Центральные опиоидергические нейроны являются важными медиаторами функции гипоталамо-гипофизарной системы. В зависимости от пептида-предшественника нейропептиды делятся на три класса: эндорфины, энкефалины и динорфины. Эндорфины (эндогенные морфины) являются продуктами расщепления гена проопиомеланокортина, который также синтезирует АКТГ и а-меланоцит-стимулирующий гормон (Taylor, 1997).
Эндорфины выполняют широкий спектр физиол. функций, включая регуляцию ТТ, настроения, репродуктивной функции, ССС и дыхательной системы и восприятия боли. Существует несколько подтипов опиоидных рецепторов. В репродуктивной оси активация μ-опиоидного рецептора β эндорфинами имеет решающее значение (Seeber, 2019).
Проопиомеланокортин преимущественно вырабатывается в передней доле гипофиза, но также в ГМ, СНС, гонадах, плаценте, ЖКТ и легких. Первичный пептид, синтезируемый по данному пути, зависит от области, в которой он вырабатывается. Напр., в ГМ преобладающими пептидами являются опиоиды, тогда как в гипофизе главным образом вырабатывается АКТГ.
Гипоталамические опиоиды играют главную роль в регуляции менструального цикла, подавляя секрецию ГнРГ гипоталамусом (Funabashi, 1994). Эстроген способствует секреции эндорфинов, которая еще больше усиливается с добавлением прогестерона (Cetel, 1985). Т.о., уровень эндорфинов повышается в фолликулярную фазу, достигает максимума в лютеиновую и заметно снижается во время менструации.
Опиоидная стимуляция и прогестерон снижают частоту импульсов ГнРГ в лютеиновой фазе, стимулируя тем самым секрецию ФСГ. Подавление секреции ГнРГ опиоидными пептидами снижается во время овуляции.
Причины такого явления изучены не до конца (King, 1984). Кроме того, при функциональной гипоталамической аменорее, обусловленной нарушением пищевого поведения, интенсивными ФН и стрессом, возрастает концентрация эндогенных опиатов.
2. Кисспептин. Гипоталамические нейроны кисспептина участвуют в процессе половой дифференцировки и инициации полового созревания и репродуктивной функции у взрослых. Эти нейроны являются частью системы KNDy. Система названа в связи с коэкспрессией кисспептина с нейрокинином В и опиоидом динорфином. Нейроны кисспептина стимулируют нейроны ГнРГ, непосредственно контролируя секрецию ГнРГ. Интересно, что одна группа нейронов кисспептина опосредует отрицательную обратную связь, тогда как др. — положительную обратную связь, наблюдаемую до овуляции (Millar, 2014; Skorupskaite, 2014; Trevisan, 2018).
В недавних исследованиях экспрессия кисспептина была выявлена во многих ненейрональных тканях репродуктивных органов. Считается, что кисспептин, вырабатываемый в яичниках, играет важную роль в развитии фолликулов, созревании яйцеклеток, овуляции и стероидогенезе (Ни, 2018). Кисспептин, вырабатываемый в матке, участвует в децидуальной трансформации эндометрия, а также он вырабатывается в плаценте на протяжении всей беременности (Сао, 2019).
Нейроны, расположенные в гипоталамусе, проецируются к нейронам кисспептина, нейронам ГнРГ и др. областям ЦНС, которые воздействуют на репродуктивную функцию. Кроме того, на активность нейронов кисспептина влияют сигналы из ЦНС и гормональная обратная связь. Т.о., модуляция кисспептина и KNDy-нейронов как таргетной терапии при нарушениях пубертата, гипоталамической аменорее, индукции овуляции и приливах является актуальной областью исследований (Yang, 2016).
Исследования также направлены на изучение кисспептина в качестве потенциального биомаркера осложнений во время беременности (Ни, 2019).
3. Другие гипоталамические нейропептиды. Секреция нейропептида Y и галанина меняется в зависимости от уровня энергозатрат, напр. при анорексии и ожирении. Оба нейропептида влияют на частоту пульсации ГнРГ и усиливают ГнРГ-индуцированную секрецию гонадотропными клетками (Lawrence, 2011; Peters, 2009). Последние исследования направлены на изучение гонадотропин-ингибирующего гормона/пептидов, связанных с ITF-амидом-3.
Данный пептид синтезируется как в гипоталамусе, так и в передней доле гипофиза, и отмечено, что его секреция происходит при подавлении гонадотропинов в ответ на воздействие стресса (Iwasa, 2018).
Также в портальную систему поступает секретируемый гипоталамическими нейронами пептид, активирующий аденилатциклазу гипофиза (РАСАР). Он связывается с рецепторами на клетках передней доли гипофиза и стимулирует секрецию гормонов, включая секрецию гонадотропина, хотя и слабее, чем ГнРГ. В свою очередь, гонадотрофы также выделяют активирующий аденилатциклазу гипофиза пептид, что свидетельствует об аутокринной/паракринной ролях этого гормона в гипофизе.
Пептид, активирующий аденилатциклазу гипофиза, модулирует экспрессию ГнРГ-рецептора, и наоборот, ГнРГ изменяет экспрессию рецепторов пептида, активирующего аденилатциклазу гипофиза, на поверхности гонадотрофов. Кроме того, ГнРГ значимо повышает экспрессию гена пептида, активирующего аденилатциклазу гипофиза, в гипофизе (Halvorson, 2014). Т.о., два вышеупомянутых важных нейропептида функционально связаны на уровне передней доли гипофиза.
4. Нутритивный статус и репродуктивная система. Начало полового созревания и менструального цикла напрямую зависит от нутритивного статуса. Важную роль играет жировая клетчатка. Она функционирует как эндокринная ткань, накапливает энергию, а также является связующим звеном между репродуктивным потенциалом и энергетическим гомеостазом. Синтезируемые жировой тканью цитокиновые факторы, называемые адипокинами, являются членами семейства цитокинов и включают лептин и адипонектин.
Пониженный уровень циркулирующего лептина и более высокий уровень адипонектина приводят к снижению энергетических ресурсов, которое может возникать при дефиците питания, расстройствах пищевого поведения или некоторых видах ФН.
По некоторым данным, снижение концентрации лептина в сыворотке крови повышает активность нейронов нейропептида У, стимулируя аппетит. И наоборот, более высокий уровень лептина в сыворотке крови повышает активность нервных сетей проопиомеланокортина и кисс-пептина. Это, в свою очередь, приводит к снижению потребления пищи и стимулирует секрецию ГнРГ, тем самым оказывая влияние на репродуктивную функцию (Chehab, 2014; Guzman, 2019). Помимо воздействия на ЦНС, лептин, вероятно, влияет на репродуктивную функцию, изменяя экспрессию рецепторов ГнРГ в гонадотропах гипофиза (Odle, 2018).
Адипонектин оказывает влияние на экспрессию кисс-пептина в гипоталамусе. Экспрессия рецепторов адипонектина наблюдается по всей гипоталамо-гипофизарно-гонадной оси, в матке, плаценте, а также выявлена у развивающегося эмбриона (Dobrzyn, 2018). Др. метаболические сигналы, опосредованные инсулином, продуцируемым ПЖЖ, и грелином, вырабатываемым желудком, также, вероятно, способствуют регуляции репродуктивной функции.
в) Гормоны передней доли гипофиза. Передняя доля гипофиза содержит пять типов клеток, продуцирующих гормоны: 1) гонадотрофы (которые продуцируют ЛГ и ФСГ); 2) лактотрофы (пролактин); 3) соматотрофы (гормон роста); 4) ТТГ и 5) АКТГ. Гонадотрофы составляют 10-15% всех гормонально активных клеток передней доли гипофиза (Childs, 1983).
Секреция гормонов гипофиза стимулируется нейроэндокринной системой гипоталамуса, за исключением пролактина, который находится под тоническим торможением. Секреция ЛГ и ФСГ регулируется высвобождением ГнРГ, который воздействует на субпопуляцию гонадотрофов передней доли гипофиза. Большинство гонадотропных клеток содержат секреторные гранулы, которые включают в себя как ЛГ, так и ФСГ. Однако значительное количество клеток являются моногормональными, т.е. секретируют только ЛГ или только ФСГ.
Кортикотропин-рилизинг-гормон является рилизинг-гормоном гипофиза, который стимулирует биосинтез и секрецию АКТГ адренокортикотрофами гипофиза. Тиреотропин-рилизинг-гормон вызывает секрецию ТТГ, также известного как тиреотропин. Разл. секретагоги гипоталамуса регулируют синтез (экспрессию) СТГ. Наконец, дофамин оказывает ингибирующее влияние на синтез пролактина. Вследствие этих регуляторных механизмов повреждение ножки гипофиза приводит к гипопитуитаризму вследствие нарушения секреции ЛГ, ФСГ, гормона роста, АКТГ и ТТГ, но к увеличению секреции пролактина.
г) Гипоталамические пептиды. Пептиды небольшого размера имеют короткий период полураспада в несколько минут из-за их быстрой деградации. Пептиды вырабатываются гипоталамусом в незначительном количестве и растворяются в периферическом кровообращении, поэтому биологически активные концентрации этих факторов ограничены передней долей гипофиза. Крайне низкие концентрации этих гормонов делают их практически недиагностируемыми в сыворотке крови. Т.о., уровни соответствующих гипофизарных факторов определяются как суррогатные маркеры.
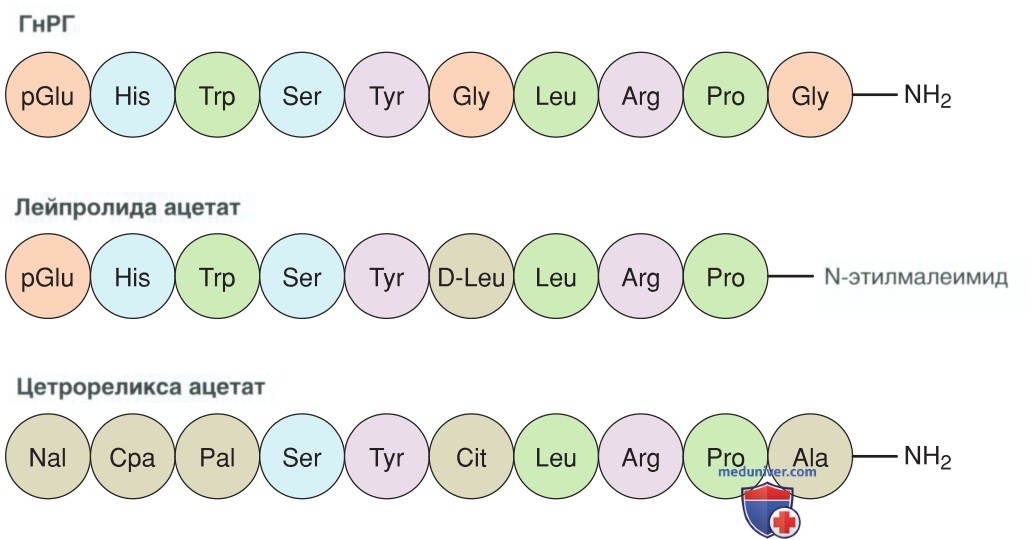
1. Гонадотропин-рилизинг-гормон. ГнРГ — декапептид с периодом полураспада <10 мин (рис. 2). Пульсация ГнРГ необходима для активации и поддержания рецепторов ГнРГ. Введение агонистов ГнРГ длительного действия используется для лечения гормонозависимых заболеваний, таких как эндометриоз, лейомиомы, преждевременное половое созревание, рак МЖ и рак предстательной железы. Агонисты ГнРГ конкурируют с эндогенным пульсирующим ГнРГ на рецепторах и тем самым угнетают секрецию гонадотропина. Это, в свою очередь, снижает уровень половых стероидов яичников в сыворотке крови.
Рисунок 2. Аминокислотный состав декапептида гонадотропин-рилизинг-гормона, агониста лейпролида ацетата и антагониста цетрореликса ацетата: Glu — глутаминовая кислота; His — гистидин; Trp — триптофан; Ser — серин; Tyr — тирозин; Gly — глицин; Leu — лейцин; Arg — аргинин; Pro — пролин; Nal— нафтилаланин; Сра — цианопропионовая аминокислота; Pal — пиридилаланин
Разл. агонисты и антагонисты ГнРГ были созданы путем введения аминокислотных модификаций в нативную последовательность ГнРГ (Padula, 2005). Изменение глицина в положении 6 привело к появлению агонистов с более длительным периодом полураспада. Напротив, изменения в положениях 1, 2 и 3 предотвращают активацию ГнРГ-рецепторов, и примерами таких антагонистов являются цетрореликс и ганиреликс. Многие из этих ЛП вводятся п/к, также существуют агонисты в форме назального спрея. В США недавно был одобрен пероральный непептидный антагонист элаголикс для лечения боли, ассоциированной с эндометриозом (Taylor, 2018).
У человека экспрессируются две формы ГнРГ: ГнРГ I и ГнРГ II (Cheng, 2005). Пептид ГнРГ II экспрессируется иначе, чем классический вариант ГнРГ, и отличается в активации рецептора (Neill, 2002). Необходимы дальнейшие исследования для определения сходства и различий в функциях двух форм рецепторов (до конца не изучены).
2. Миграция гонадотропин-рилизинг-гормона нейронов. Большая часть гипоталамических нейронов формируется в ЦНС, но ГнРГ-содержащие нейроны имеют уникальное эмбриологическое происхождение. Предшественники ГнРГ-нейронов возникают в медиальной части обонятельной плакоды и мигрируют по вомероназальному нерву в гипоталамус. Ряд растворимых факторов регулируют миграцию ГнРГ-нейронов. Эти факторы включают сигнальные молекулы, такие как ГАМК, молекулы адгезии и факторы роста (Wierman, 2011).
Нарушение миграции вследствие разл. генетических дефектов в этих сигнальных молекулах может привести к синдрому Каллмана, который описан в отдельной статье на сайте - просим Вас пользоваться формой поиска по сайту выше.
Нейроны ГнРГ расположены преимущественно в аркуатных ядрах, откуда ГнРГ транспортируется по аксональному пути, именуемому тубероинфундибулярным трактом к срединному возвышению. Затем ГнРГ поступает в портальную систему и непосредственно в переднюю долю гипофиза, стимулирует биосинтез и секрецию гонадотропинов. У взрослого человека удивительно мало ГнРГ-нейронов, и только несколько тысяч клеток расположены внутри дугообразного ядра.
Происхождение нейронов ГнРГ из обонятельной области и клеток носового эпителия предполагает связь между репродукцией и обонятельными сигналами. В-ва, выделяемые одним представителем вида, которые оказывают влияние на др. представителей того же вида, называются феромонами. Феромоны, вырабатывающиеся в подмышечных впадинах у женщин в позднюю фолликулярную фазу, ускоряют овуляторный пик ЛГ, что приводит к сокращению менструального цикла.
Секреция феромонов в лютеиновой фазе оказывает противоположный эффект. Т.о., феромоны м.б. одним из механизмов, влияющим на синхронизацию менструальных циклов у женщин, которые часто находятся вместе (Stern, 1998).
Часть ГнРГ-нейронов проецируются (посылают проекции) в др. области ЦНС, включая лимбическую. Эти проекции не влияют на секрецию гонадотропинов, но, вероятно, они могут играть определенную роль в модуляции репродуктивного поведения (Nakai, 1978; Silverman, 1987).
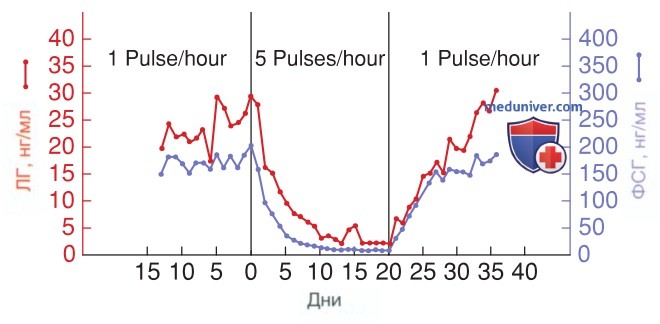
3. Импульсный паттерн секреции гонадотропин-рилизинг-гормона. В 1974 г. Кнобил провел ряд экспериментов, доказав, что для достижения устойчивой секреции гонадотропинов необходимо импульсное выделение ГнРГ в гонадотропы гипофиза. Как показано на рис. 3, непрерывное воздействие ГнРГ быстро снижает секрецию как ЛГ, так и ФСГ. Этот эффект легко обратить вспять с помощью импульсной стимуляции (Knobil, 1980).
Рисунок 3. Изменение уровня лютеинизирующего гормона и фолликулостимулирующего гормона при изменении импульсной секреции гонадотропин-рилизинг-гормона
По сравнению с лютеиновой фазой выброс ГнРГ в фолликулярной фазе характеризуется более высокой частотой импульсов и меньшей их амплитудой. Более высокая частота импульсов преимущественно стимулирует секрецию ЛГ, в то время как более низкая способствует секреции ФСГ (Thompson, 2014). Т.о., изменения частоты импульсов ГнРГ влияют на абсолютные уровни и соотношения концентраций ЛГ и ФСГ.
Импульсная активность считалась неотъемлемым свойством ГнРГ-нейронов. Согласно последним данным, нейроны кисспептина, вероятно, оказывают влияние на импульсную секрецию ГнРГ (Terasawa, 2019). На частоту и амплитуду импульсной выработки ГнРГ влияет ряд нейромедиаторов и циркулирующих гормонов. При проведении экспериментов на животных моделях было выявлено, что эстрогены увеличивают частоту импульсов ГнРГ и, следовательно, приводят к увеличению уровня ЛГ относительно ФСГ. Напротив, прогестерон снижает частоту импульсов ГнРГ.
Увеличение концентрации прогестерона во время лютеиновой фазы приводит к секреции преимущественно ФСГ, что наблюдается к концу фазы. Повышение уровня ФСГ имеет решающее значение для стимуляции рекрутирования фолликулов.
д) Другие оси гипоталамо-гипофизарной системы:
1. Дофамин и пролактин. В отличие от др. гормонов передней доли гипофиза, выработка пролактина в основном регулируется посредством ингибирования, в частности дофамином (табл. 4). Дофаминсодержащие нервные волокна возникают главным образом в дугообразном ядре гипоталамуса и проецируются на срединное возвышение, где дофамин попадает в сосуды воротной вены. Пролактин-рилизинг-факторы, хотя и менее активные, включают тиреотропин-рилизинг-гормон, вазопрессин, вазоактивный интестинальный пептид, эндогенные опиоиды и ацетилхолин.
Существует пять форм дофаминовых рецепторов, которые подразделяются на две группы: D1 и D2. Клетки передней доли гипофиза в основном экспрессируют подтипы D2. Разработка D2-специфических лигандов привела к улучшению эффективности и переносимости медикаментозного лечения пролактином. Напр., агонист дофамина каберголин является D2-специфическим лигандом, тогда как бромокриптин неселективен.
2. Тиреотропин-рилизинг-гормон. Как следует из его названия, тиреотропин-рилизинг-гормон стимулирует секрецию ТТГ из субпопуляции тиреотрофов передней доли гипофиза. Следует отметить, что тиреотропин-рилизинг-гормон также является активным пролактин-рилизинг-фактором, что объясняет связь между гипотиреозом и вторичной гиперпролактинемией (Messini, 2010).
ТТГ связывается со специфическими рецепторами на плазматической мембране клеток ЩЖ. Это стимулирует биосинтез тиреоидных гормонов за счет увеличения размеров ЩЖ и ее кровоснабжения. Тиреоидный гормон тормозит секрецию тиреотропин-рилизинг-гормона и ТТГ по принципу отрицательной обратной связи.
3. Кортикотропин-рилизинг-гормон. Это основной гормон гипоталамуса, стимулирующий синтез и секрецию АКТЕ Кортикотропин-рилизинг-гормон синтезируется в нескольких участках гипоталамуса и др. областях ЦНС. Выброс кортикотропин-рилизинг-гормона стимулируется катехоламинергическими агентами и замедляется под влиянием эндогенных опиоидов.
Кортикотропин-рилизинг-гормон связывается с рецепторами кортикотропин-рилизинг-гормона в передней доле гипофиза, стимулируя биосинтез и секрецию АКТЕ В свою очередь, АКТЕ стимулирует выработку глюкокортикоидов в пучковой зоне надпочечников и андрогенов в сетчатой зоне. Секреция кортикотропин-рилизинг-гормона регулируется кортизолом, вырабатываемым надпочечниками, посредством отрицательной обратной связи. Выработка минералокортикоидов в клубочковой зоне регулируется в первую очередь РААС. Изменения в пути кортикотропин-рилизинг-гормон-АКТГ не приводят к возникновению электролитных нарушений.
Считается, что пути кортикотропин-рилизинг-гормона опосредуют многие стрессовые реакции (Kalantaridou, 2004). Было обнаружено, что у женщин с гипоталамической аменореей уровень кортикотропин-рилизинг-гормона повышен. При его повышении происходит ингибирование секреции ГнРГ в гипоталамусе путем прямого воздействия, а также увеличение концентрации опиоидов. Этот функциональный путь частично объясняет связь между гиперкортицизмом и нарушениями менструального цикла. Однако, согласно последним данным, эта связь м.б. существенно более сложной и включать в себя несколько нейронных сетей и нейромедиаторов (McCosh, 2019).
4. Гормон, высвобождающий соматотропный гормон. Секреция СТГ соматотропами гипофиза стимулируется соматотропин-рилизинг-гормоном и ингибируется соматостатином. Соматотропин-рилизинг-гормон секретируется в основном гипоталамусом, но небольшое количество вырабатывается иммунными клетками и клетками плаценты. Напротив, соматостатин в большом количестве содержится в ЦНС и в плаценте, ПЖЖ и ЖКТ.
Как и в случае ГнРГ, соматотропин-рилизинг-гормон зависит от импульсной секреции для поддержания физиол. действия. ФН, стресс, сон и гипогликемия стимулируют секрецию СТГ, в то время как свободные жирные кислоты и др. факторы, связанные с ожирением, тормозят его выработку. Эстроген, тестостерон и тиреоидный гормон также влияют на секрецию гормона роста.
СТГ стимулирует рост костной и мышечной тканей (скелета и мышц), регулирует липолиз и способствует усвоению аминокислот клетками. Установлено, что гормон роста индуцирует инсулинорезистентность, что может приводить к развитию СД в будущем. Ростостимулирующие воздействия СТГ опосредуются через инсулиноподобные факторы роста-1 и 2, которые синтезируются главным образом в печени. Многие ткани-мишени, на которые они оказывают воздействие, также синтезируют инсулиноподобные факторы роста. В яичнике инсулиноподобные факторы роста-1 и 2 стимулируют пролиферацию гранулезных клеток и стероидогенез во время фолликулогенеза (Silva, 2009).
Инсулиноподобные факторы роста также подавляют секрецию СТГ посредством механизма отрицательной обратной связи.
е) Пептиды задней доли гипофиза. Нейроны, которые проецируются в заднюю долю гипофиза, синтезируют и секретируют циклические пептиды, состоящие из 9 аминокислот, — окситоцин и аргинин-вазопрессин. Предшественники этих пептидов вырабатываются в теле нейрональной клетки и транспортируются вниз по аксону в секреторные гранулы. Во время транспортировки молекулы-предшественники расщепляются на зрелые пептиды и белок-переносчик — нейрофизин (Verbalis, 1983).
Активация нейронов генерирует потенциал действия, который приводит к притоку кальция и секреции содержимого гранул в периваскулярное пространство. Секретируемые пептиды через кровеносные сосуды распределяются по периферическому кровообращению.
Из двух вышеупомянутых пептидов задней доли гипофиза окситоцин играет важную роль как в процессе родов, так и в период лактации (Kiss, 2005). Роль окситоцина в инициации родов спорна, т.к. его уровень в сыворотке крови остается неизменным вплоть до второго периода родов (Blanks, 2003). Однако интересно, что перед родами отмечается большая экспрессия окситоциновых рецепторов в миометрии и децидуальной ткани, преимущественно за счет повышения уровня эстрогенов.
После начала родов окситоцин является основным медиатором сократительной активности миометрия. Стимуляция шейки матки и стенок влагалища приводит к выработке окситоцина в задней доле гипофиза в процессе рефлекса Фергюсона. Способность окситоцина индуцировать сокращения матки используется в клинической практике для стимуляции родовой деятельности.
Выработка окситоцина также способствует расширению стенок влагалища, как при половом акте. На основании этого можно предположить, что окситоцин вызывает ритмичные сокращения матки и маточных труб, которые способствуют доставке сперматозоида к яйцеклетке. Окситоцин также может влиять на оргазм и эякуляцию.
Во время лактации пролактин имеет особое значение для выработки молока в альвеолах МЖ. Железистые клетки альвеол окружены миоэпителиальными клетками. Акт сосания вызывает нервные импульсы от механорецепторов в соске и ареоле, которые повышают гипоталамическую нейрональную активность. Последующая выработка окситоцина приводит к сокращению миоэпителиальных клеток и выделению молока из альвеол в протоки и синусы (Crowley, 1992). Др. стимулы, такие как вид ребенка, его крик или запах, а также сексуальное возбуждение могут оказывать аналогичный эффект.
Окситоцин также вырабатывается в передней доле гипофиза, плаценте, маточных трубах, гонадах и желтом теле яичника (Williams, 1990). Его функция в этих тканях неясна.