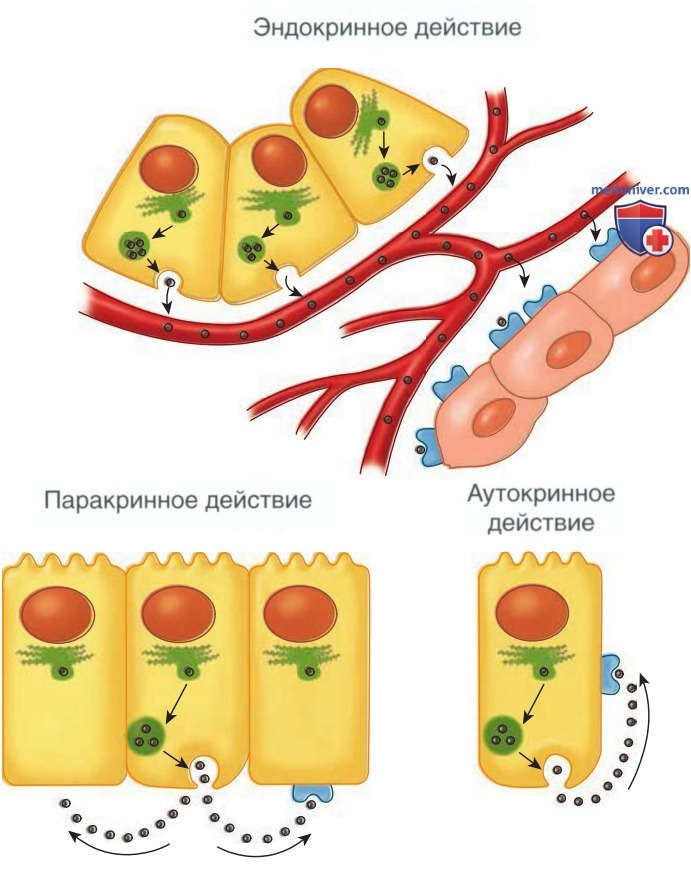
Репродуктивная эндокринология — раздел науки, изучающий нейроэндокринные механизмы регуляции половых гормонов, которые синтезируются тканями репродуктивной системы и/или оказывают на них воздействие. К таким тканям относятся гипоталамус, передняя доля гипофиза, яичники, эндометрий и плацента. Традиционно гормоны описываются как продукты деятельности специализированных клеток, которые поступают в периферическое кровообращение и влияют на ткани-мишени (рис. 1).
Рисунок 1. Разл. типы коммуникации гормонов. Эндокринная: гормоны проходят через кровяное русло и достигают клетки-мишени. Паракринная: гормоны проходят через внеклеточное пространство и достигают соседних клеток-мишеней. Аутокринная: гормоны влияют на те клетки, которые их продуцируют, и не попадают в кровяное русло
Данный процесс называется эндокринной секрецией. В репродуктивной физиологии существуют дополнительные формы межклеточных коммуникаций. Паракринная коммуникация происходит внутри яичника. При этом типе коммуникации хим. сигнальные механизмы действуют только на соседние клетки. При аутокринной коммуникации клетка высвобождает в-ва, влияющие на ее собственную функцию. Выработка в-в внутри клетки, которые воздействует на эту клетку до секреции, называется интракринным механизмом.
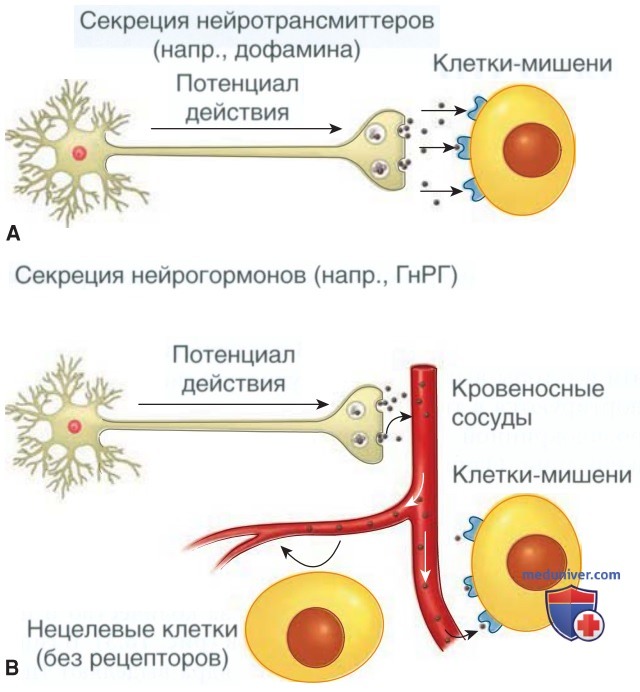
Нейротрансмиттеры поступают в синапс — небольшое внеклеточное пространство и связываются с дендритами второго нейрона (рис. 2). В альтернативных случаях эти факторы выделяются в сосудистую систему и транспортируются в ткани, где они участвуют в процессе нейроэндокринной секреции или нейроэндокринной сигнализации. Одним из примеров является секреция ГнРГ в портальную сосудистую систему, который воздействует на гонадотрофы передней доли гипофиза.
Рисунок 2. Типы секреции нейромедиаторов: А — классический тип высвобождения и связывания нейромедиаторов. Передача потенциала действия вниз по нервному аксону приводит к высвобождению нейромедиаторов, которые перемещаются через синаптическую щель и достигают клетки-мишени; В — секреция нейрогормонов. Потенциал действия приводит к высвобождению нейромедиаторов. В этом случае нейромедиаторы поступают в кровеносное русло и достигают органов-мишеней
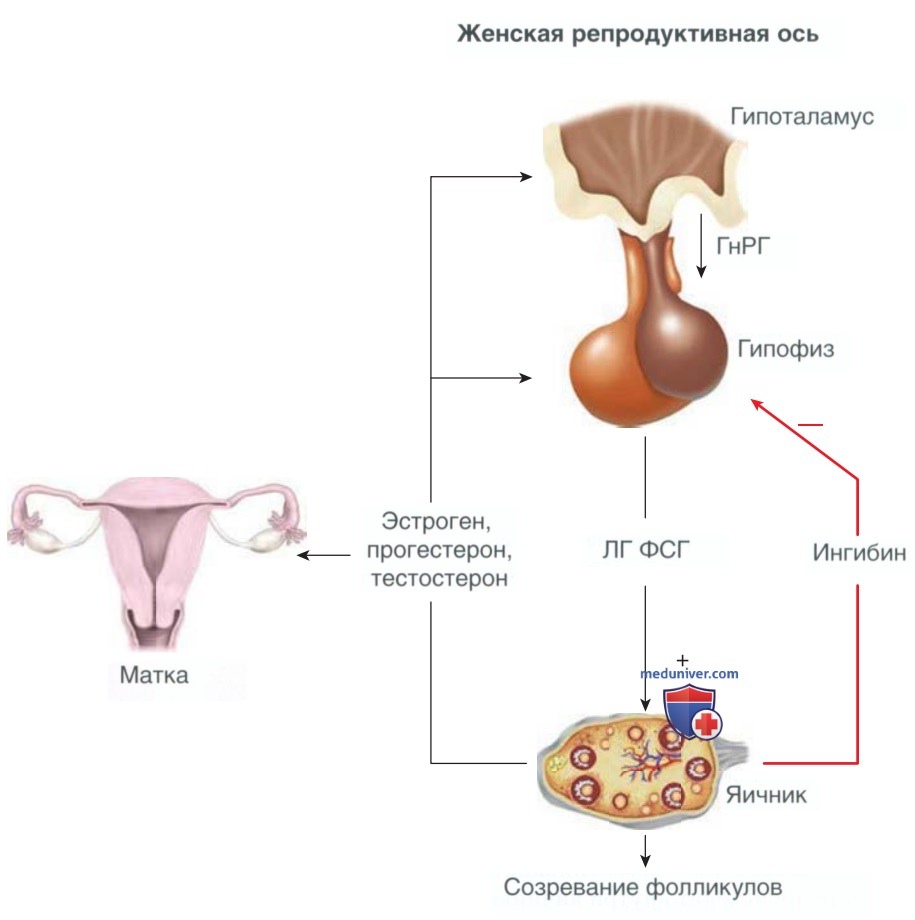
Для нормальной работы репродуктивной функции необходима регуляция гипоталамо-гипофизарно-яичниковой системы, которая отвечает за количество и цикличность синтеза половых гормонов (рис. 3). В гипоталамусе определенные центры или ядра выделяют ГнРГ в виде импульсов. Декапептид связывается с рецепторами на поверхности гонадотропных клеток передней доли гипофиза. В ответ гонадотрофы выделяют в периферическое кровообращение гликопротеиновые гонадотропины, а именно ЛГ и ФСГ.
Рисунок 3. Петли положительной и отрицательной обратных связей, наблюдаемые на оси гипоталамус-гипофиз-яичники. Импульсная секреция гонадотропин-рилизинг-гормона приводит к высвобождению лютеинизирующего гормона и фолликулостимулирующего гормона из передней доли гипофиза. Лютеинизирующий гормон и фолликулостимулирующий гормон приводят к созреванию фолликулов, овуляции и выработке половых стероидных гормонов (эстрогена, прогестерона и тестостерона). Повышение уровня гормонов в сыворотке крови приводит к ингибированию по принципу отрицательной обратной связи, уменьшая скорость секреции гонадотропин-рилизинг-гормона и гонадотропина. Стероидные половые гормоны оказывают разное воздействие на эндометрий и миометрий. Ингибин, вырабатываемый в яичнике, снижает секрецию гонадотропина
Внутри яичника ЛГ и ФСГ связываются с текаклетками и гранулезными клетками, стимулируя фолликулогенез и секрецию яичниками стероидных гормонов (эстрогенов, прогестерона и андрогенов), гонадных пептидов (активина, ингибина и фоллистатина) и факторов роста. Помимо др. функций, данные факторы воздействуют на гипоталамус и гипофиз по принципу обратной связи, подавляя или в середине цикла усиливая секрецию ГнРГ и гонадотропина. Стероидные гормоны яичника также способствуют изменениям в эндометрии, необходимым для успешной имплантации.
В широком смысле гормоны можно разделить на стероиды или пептиды. Они отличаются способом биосинтеза и механизмом действия. Гормональные рецепторы можно разделить на две группы. Первые — рецепторы, присутствующие на поверхности клеток, которые связывают водорастворимые гормоны — пептиды. Вторые являются преимущественно в/клеточными и связывают липофильные гормоны, такие как стероиды.
Как правило, гормоны присутствуют в сыворотке крови и тканях в очень низких концентрациях, в связи с этим рецепторы должны обладать как высокой аффинностью, так и высокой специфичностью, чтобы их лиганд вызывал правильный биологический ответ.
а) Пептидные гормоны: лютеинизирующий гормон, фолликулостимулирующий гормон и хорионический гонадотропин человека. Гонадотропины ЛГ и ФСГ биосинтезируются и секретируются гонадотропными клетками передней доли гипофиза. Данные гормоны играют важную роль в стимуляции стероидогенеза яичников, развитии фолликулов и овуляции. Близкородственный им пептид, ХГЧ, вырабатывается клетками синцитиотрофобласта плаценты и имеет большое значение для поддержания беременности.
ЛГ, ФСГ и ХГЧ являются гетеродимерами, состоящими из общей α-субъединицы гликопротеина, связанной с уникальной β-субъединицей. β-Субъединица обеспечивает функциональную специфичность. Гликопротеиновые α- и β-субъединицы могут циркулировать в несвязанной форме, однако данные «свободные» субъединицы не обладают биологической активностью. Тем не менее их количество определяется при проведении скрининга на аденому гипофиза или с целью диагностики беременности.
β-Субъединицы ЛГ и ХГЧ кодируются двумя отдельными генами в общей группе, называемой кластером ЛГ/ХГЧ. Аминокислотная последовательность β-субъединиц ЛГ и ХГЧ совпадает на ~80%, β-субъединица ХГЧ содержит дополнительное ответвление на карбоксильном конце, состоящее из 24 аминокислот. Наличие дополнительных этих аминокислот позволило разработать высокоспецифичные анализы, с помощью которых можно отличать ЛГ от ХГЧ.
В клетках гипофиза α-субъединица гликопротеина взаимодействует со специфической β-субъединицей ТТГ, образуя ТТГ. Сходство между ТТГ и ХГЧ имеет клиническое значение. Напр., при пузырном заносе диагностируется очень высокий уровень ХГЧ, который может связываться с рецепторами ТТГ, что в результате приводит к гипертиреозу (Walkington, 2011).
б) Хорионический гонадотропин человека (ХГЧ). ХГЧ — гликозилированный пептидный гормон, который вырабатывается синцитиотрофобластом плаценты. Степень и тип гликозилированных фрагментов, прикрепленных к пептиду ХГЧ, могут варьировать и указывать на срок беременности, состояние плаценты и наличие патологии (Fournier, 2015). Одним из примеров является гипергликозилированный ХГЧ, который чаще встречается при гестационной трофобластической неоплазии.
ХГЧ м.б. обнаружен в сыворотке крови уже через 7-9 сут после овуляторного пика ЛГ. На ранних сроках беременности уровень ХГЧ стремительно повышается, удваиваясь примерно каждые 2 сут, достигая максимума в ~100 000 мМЕ/мл в течение I триместра беременности. В начале II триместра происходит относительно резкое снижение концентрации ХГЧ, которая сохраняется на протяжении оставшегося периода беременности.
На ранних сроках беременности ХГЧ связывается с рецепторами ЛГ/ХГЧ на клетках желтого тела и стимулирует стероидогенез в яичниках. Секреция ХГЧ имеет решающее значение для поддержания интактности эндометрия и матки, выработки желтым телом стероидов на ранних сроках беременности до достижения нормальной стероидогенной способности плаценты. Прекращение выработки эстрогенов и прогестерона яичниками и начало их выработки плацентой обычно называют лютеоплацентарным сдвигом. Помимо влияния на функцию яичников, ХГЧ оказывает аутокринное/паракринное действие в плаценте, способствуя образованию синцитиотрофобласта, инвазии трофобласта и ангиогенезу.
Поскольку плацента является основным источником выработки ХГЧ, измерение его уровня в плазме крови является эффективным инструментом скрининга беременностей соответственно морфофункциональным изменениям плаценты. Относительно повышенные уровни ХГЧ наблюдаются при многоплодных беременностях и при синдроме Дауна у плода. Выраженное аномальное повышение уровня ХГЧ чаще всего бывает при гестационной трофобластической неоплазии.
Сниженный уровень ХГЧ наблюдается при нарушениях плацентации, внематочной беременности или самопроизвольном выкидыше. В последнее время исследования направлены на разработку алгоритмов на основе последовательного измерения уровня ХГЧ и данных УЗИ для точного определения статуса беременности неизвестной локализации (Zee, 2014). Время удвоения уровня ХГЧ при нормальной беременности является относительно надежным показателем.
ХГЧ также секретируется нетрофобластными новообразованиями и является важным опухолевым маркером. Эктопическая (неплацентарная) продукция ХГЧ, либо интактного димера, либо β-субъединицы, часто связана с герминогенной опухолью и также обнаруживалась при опухолях, возникающих из эпителия слизистой оболочки шейки матки, мочевого пузыря, легких, ЖКТ и носоглотки. Было высказано предположение, что ХГЧ замедляет апоптоз в опухолях и тем самым обеспечивает быстрый рост.
ХГЧ секретируется не только синцитиотрофобластом и, вероятно, выполняет др. функции (Cole, 2010). Напр., цитотрофобласты синтезируют гипергликозилированный вариант ХГЧ, который может оказаться чувствительным маркером ранних сроков беременности (Chuan, 2014). Гонадотрофы гипофиза также производят небольшое количество ХГЧ. Его концентрация повышается у женщин в постменопаузе, что м.б. редкой причиной л/п-теста на ХГЧ в этой возрастной группе (Cole, 2008).
в) Стероидные гормоны:
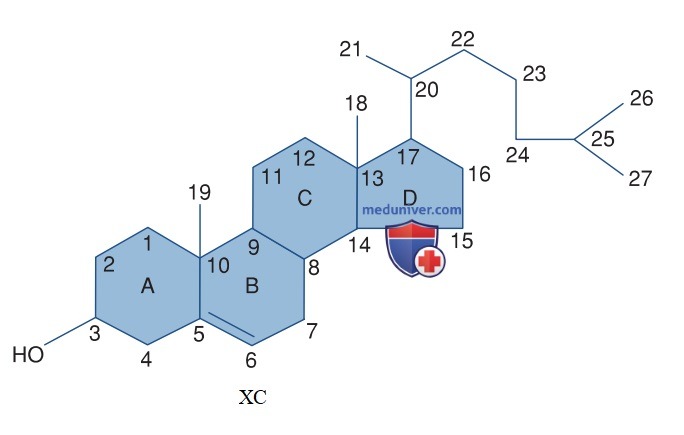
1. Классификация. Половые стероиды делятся на три группы в зависимости от количества атомов углерода, которые они содержат. Каждый углерод в этой структуре обозначается числом, а каждое кольцо — буквой (рис. 4). Ряд из 21 атома углерода включает прогестагены, глюкокортикоиды и минералокортикоиды. Андрогены содержат 19 атомов углерода, а эстрогены — 18.
Рисунок 4. Хим. строение холестерина, который является общим предшественником в биосинтезе половых стероидов. Все половые стероиды содержат основную молекулу циклопентанофенантрена, которая состоит из трех 6-углеродных колец и одного 5-углеродного кольца
Стероидам дают научные названия в соответствии с общепринятой конвенцией. Функциональные группы под плоскостью молекулы обозначаются символом α, а группы над плоскостью молекулы — символом β. Символ Д указывает на двойную связь. Эти стероиды с двойной связью между атомами углерода 5 и 6 называются стероидами Δ5 и включают прегненолон, 17-гидроксипрегненолон и дегидроэпиандростерон. Стероиды, которые имеют двойную связь между углеродами 4 и 5, называются стероидами Δ4 и включают прогестерон, 17-гидроксипрогестерон, андростендион, тестостерон, минералокортикоиды и глюкокортикоиды.
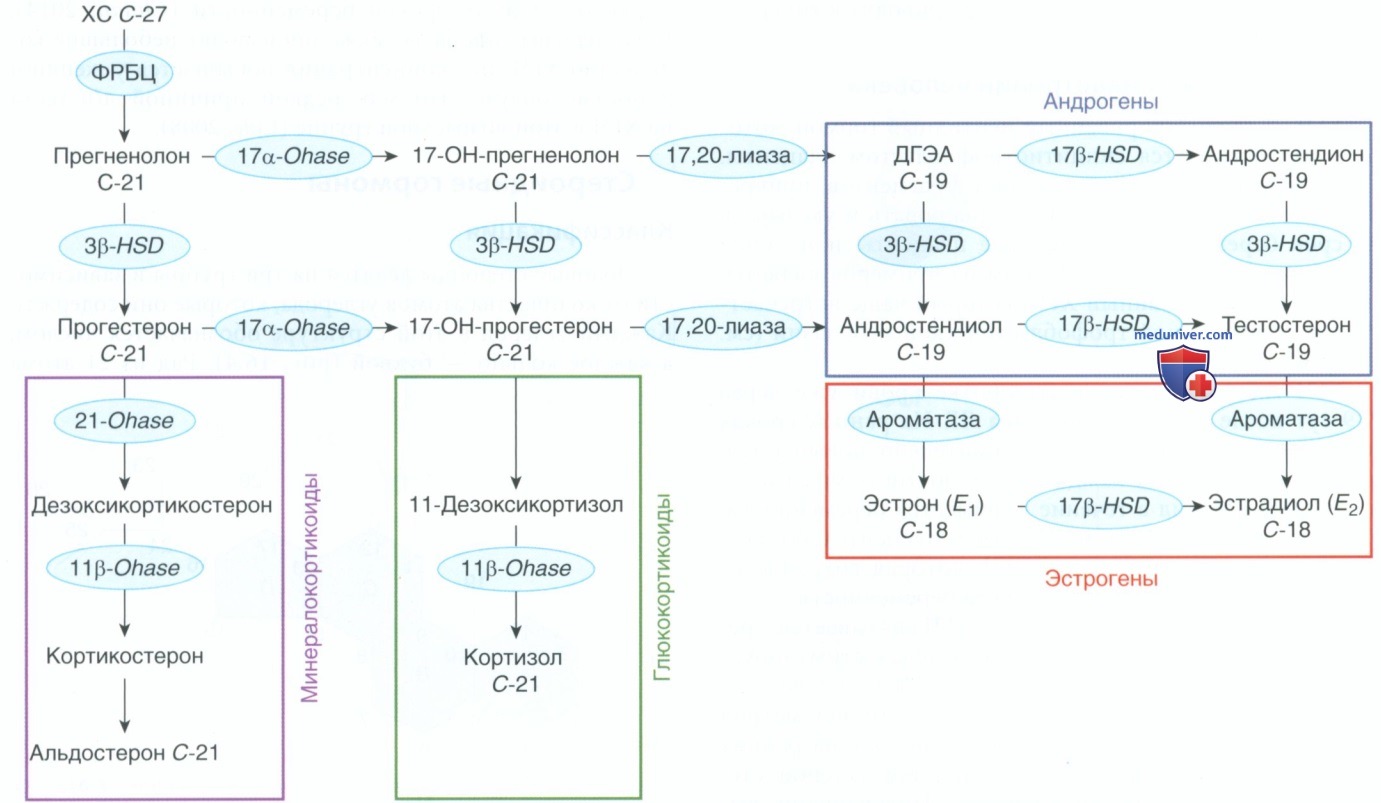
2. Стероидогенез. Половые стероидные гормоны синтезируются в гонадах, надпочечниках и плаценте. ХС является основным строительным материалом для продукции гормонов. Все ткани, вырабатывающие стероиды, за исключением плаценты, способны синтезировать ХС из двухуглеродного предшественника-ацетата. Синтез стероидных гормонов катализируется по меньшей мере 17 ферментами и происходит преимущественно в митохондриях и гладком эндоплазматическом ретикулуме, базирующихся в стероидогенных клетках (Mason, 2002). Эти ферменты являются членами суперсемейства цитохрома Р450. Т.о., название генов, кодирующих эти ферменты, начинается с CYP.
Стероидогенные ферменты катализируют четыре основные модификации структуры стероидов: 1) расщепление боковых цепей (десмолазная реакция); 2) превращение гидроксильных групп в кетоны (дегидрогеназные реакции); 3) добавление гидроксильной группы (реакция гидроксилирования) и 4) удаление или добавление водорода для создания или уменьшения двойной связи (лиазная реакция) (табл. 1). Путь биосинтеза стероидов в упрощенном виде показан на рис. 5. Этот путь идентичен во всех стероидогенных тканях.
Рисунок 5. Стадии стероидогенеза. Ферменты обведены синим цветом. Обозначение С18, С19 или С21 под половым стероидом отражает количество атомов углерода, которые он содержит. В цветных рамках отмечены гормоны по группам: 3β-HSD — 3β-гидроксистероиддегидрогеназа; 11β-OHase — 11β-гидроксилаза; 17α-OHase — 17α-гидроксилаза; 17β-HSD — 17β-гидроксистероиддегидрогеназа; 21OHase — 21-гидроксилаза; ДГЭА — дегидроэпиандростерон; ФРБЦ — фермент расщепления боковой цепи
Однако распределение продуктов, синтезируемых отдельными тканями, определяется наличием необходимых ферментов. Напр., отсутствуют ферменты 21-гидроксилазы и 11β-гидроксилазы, вследствие чего яичники не способны вырабатывать глюкокортикоиды. Следует отметить, что многие стероидогенные ферменты существуют в виде нескольких изоформ, для каждой из которых характерны разл. предшественники и направленная активность. Установлено, что помимо классического пути, описанного на рис. 5, стероиды могут синтезироваться иными специфическими путями (Auchus, 2009).
Среди специфических гормонов эстрогены синтезируются путем ароматизации андрогенов С19 ферментом ароматазой. Этот энзим представляет собой фермент цитохрома Р450, кодируемый геном CYP19. Помимо яичников, ароматаза в значительном количестве вырабатывается в жировой ткани, коже и ГМ (Boon, 2010). Важно отметить, что достаточное количество эстрогена синтезируется путем периферической ароматизации, вызывая маточное кровотечение в постменопаузальном периоде, особенно у женщин с избыточной МТ или ожирением.
Циркулирующие эстрогены у женщин репродуктивного возраста включают эстрадиол (Е2), эстрон (Е1) и эстриол (Е3). Эстрадиол является основным эстрогеном, вырабатываемым яичниками в репродуктивном возрасте как путем прямого синтеза в гранулезных клетках созревающих фолликулов, так и путем преобразования менее активного эстрона. Эстрон, эстроген, преимущественно синтезирующийся в период менопаузы, секретируется главным образом яичниками. А также эстриол, который преобладает во время беременности, выделяется главным образом плацентой. Однако и эстрон, и эстриол м.б. преобразованы из андростендиона в периферических тканях.
В процессе синтеза андрогенов фермент 5α-редуктаза преобразует тестостерон в ДГТ, более активный андроген. Под влиянием ДГТ пушковые волосы преобразуются в терминальные. Т.о., ЛП — антагонисты 5α-редуктазы, часто эффективны при лечении гирсутизма (Stout, 2010). Данный фермент существует в двух формах, каждая из которых кодируется отдельным геном. Фермент 1-го типа содержится в коже, ГМ, печени и почках, в то время как фермент 2-го типа синтезируется преимущественно мужскими половыми органами (Russell, 1994).
Яичники также вырабатывают андрогены текаклетками в ответ на стимуляцию ЛГ. Два основных гормона — относительно «слабые» андрогены: андростендион и дегидроэпиандростерон, а также синтезируется небольшое количество тестостерона. Хотя кора надпочечников в основном секретирует минерало- и глюкокортикоиды, там же вырабатывается 50% суточного количества андростендиона и дегидроэпиандростерона и практически вся сульфатированная форма дегидроэпиандростерона. У женщин 25% циркулирующего в крови тестостерона вырабатывается яичниками, 25% — надпочечниками, а остальные 50% — при преобразовании андростендиона в тестостерон (табл. 2) (Silva, 1987).
Надпочечник взрослого человека подразделяется на три зоны. Каждая зона вырабатывает разл. набор стероидогенных ферментов и в результате синтезирует разл. гормоны. В клубочковой зоне отсутствует 17α-гидроксилаза, но она содержит большое количество альдостеронсинтазы (Р450) и поэтому производит минералокортикоиды. Пучковая сетчатая зона экспрессирует ген 17α-гидроксилазы, синтезирует глюкокортикоиды и андрогены соответственно.
3. Транспорт стероидных гормонов в системе кровообращения. Большинство стероидов в периферическом кровообращении связываются с белками-носителями. Белки м.б. специфическими, такими как глобулин, связывающий половые гормоны, глобулин, связывающий гормоны ЩЖ, или глобулин, связывающий глюкокортикоиды, или неспецифическими, напр. альбумин. Только 1-2% циркулирующих андрогенов и эстрогенов являются несвязанными, или свободными.
Только свободные стероидные фракции могут быть биологически активными. Однако низкое сродство (относительная аффинность) альбумина к половым стероидам позволяет стероидам, связанным с этим белком, проявлять некоторую активность. Количество свободного, биологически активного гормона обратно пропорционально количеству связанного гормона, а количество связанного гормона соответствует уровню белка-носителя. В результате небольшие изменения в биосинтезе белка-носителя могут существенно изменить активность стероидных гормонов.
Глобулин, связывающий половые гормоны, в виде гомодимера связывается с молекулой стероидного гормона. Этот связывающий белок синтезируется главным образом в печени, хотя он также был обнаружен в ГМ, плаценте, эндометрии и гонадах. Уровень связывающего половые гормоны глобулина повышается при гипертиреозе, беременности и экзогенном применении эстрогенов. Напротив, андрогены, прогестины, гормон роста, инсулин и глюкокортикоиды снижают уровень связывающего половые гормоны глобулина.
Увеличение жировой ткани, особенно в абдоминальной области, может значительно замедлить выработку глобулина, связывающего половые гормоны. В свою очередь, это приводит к снижению уровня связанных гормонов и повышению уровня активных гормонов (Hammond, 2012).
Из стероидных гормонов чаще всего определяют уровень свободного тестостерона. Наиболее точные анализы выполняются только несколькими коммерческими лабораториями (Rosner, 2007). Хотя масс-спектрометрия считается предпочтительным методом исследования, она не получила широкого распространения из-за сложности и высокой стоимости метода (Kanakis, 2019). Результаты более доступных методов диагностики являются относительно неточными. Необходимо с особым вниманием подходить к интерпретации полученных данных.
В клинической практике обычно нет необходимости определения тестостерона свободного, более информативна оценка общего тестостерона. Уровень тестостерона определяют при подозрении на СПКЯ для исключения андрогенпродуцирующей опухоли, для которой патогномонично повышение уровня общего тестостерона. При диагнозе СПКЯ уровень общего тестостерона м.б. нормальным или повышенным.
Поскольку тестостерон снижает уровень связывающего половые гормоны глобулина, пациенты с нормальным уровнем общего тестостерона, но с клиническими признаками гиперандрогении (гирсутизм и/или акне) имеют либо повышенный уровень свободного тестостерона, либо повышенную чувствительность волосяных фолликулов и сальных желез.
Стероиды метаболизируются главным образом в печени и в меньшей степени в почках и слизистой оболочке кишечника. Гидроксилирование эстрадиола приводит к образованию эстрона или катехол-эстрогенов. Затем эстрогены соединяются с глюкуронидами или сульфатами, образуя водорастворимые соединения, которые выводятся с мочой. Соответственно, введение некоторых стероидных гормонов м.б. противопоказано пациентам с активными заболеваниями печени или почек.