Врожденная иммунная система - кратко с точки зрения внутренних болезней
Иммунная система развилась в ходе эволюции для выявления и уничтожения патогенных микроорганизмов при минимальном повреждении тканей собственного организма. Хотя издревле было известно, что выздоровление от инфекционного заболевания часто приводит к обретению защиты от него, существование иммунной системы как функционального объекта не признавалось до конца XIX в. Совсем недавно стало ясно, что иммунная система не только защищает организм от инфекции, но и регулирует восстановление тканей после травмы, а при нарушении ее функции развиваются реакции, которые могут привести к аутоиммунным и аутовоспалительным заболеваниям.
Дисфункция или дефицит иммунного ответа способны вызвать широкий спектр заболеваний, которые могут поражать любую систему органов.
Цель статей раздела внутренние болезни сайта — дать общее представление об иммунной системе и ее участии в развитии различных заболеваний у человека, а также о возможностях воздействия на нее в целях достижения терапевтического эффекта. После обзора основных компонентов иммунного ответа следуют разделы, иллюстрирующие клиническую картину наиболее распространенных форм иммунной дисфункции: иммунодефицита, воспаления, аутоиммунных реакций и аллергии. Более подробное обсуждение отдельных заболеваний приводится в соответствующих статьях на сайте (просим вас пользоваться формой поиска по сайту выше).
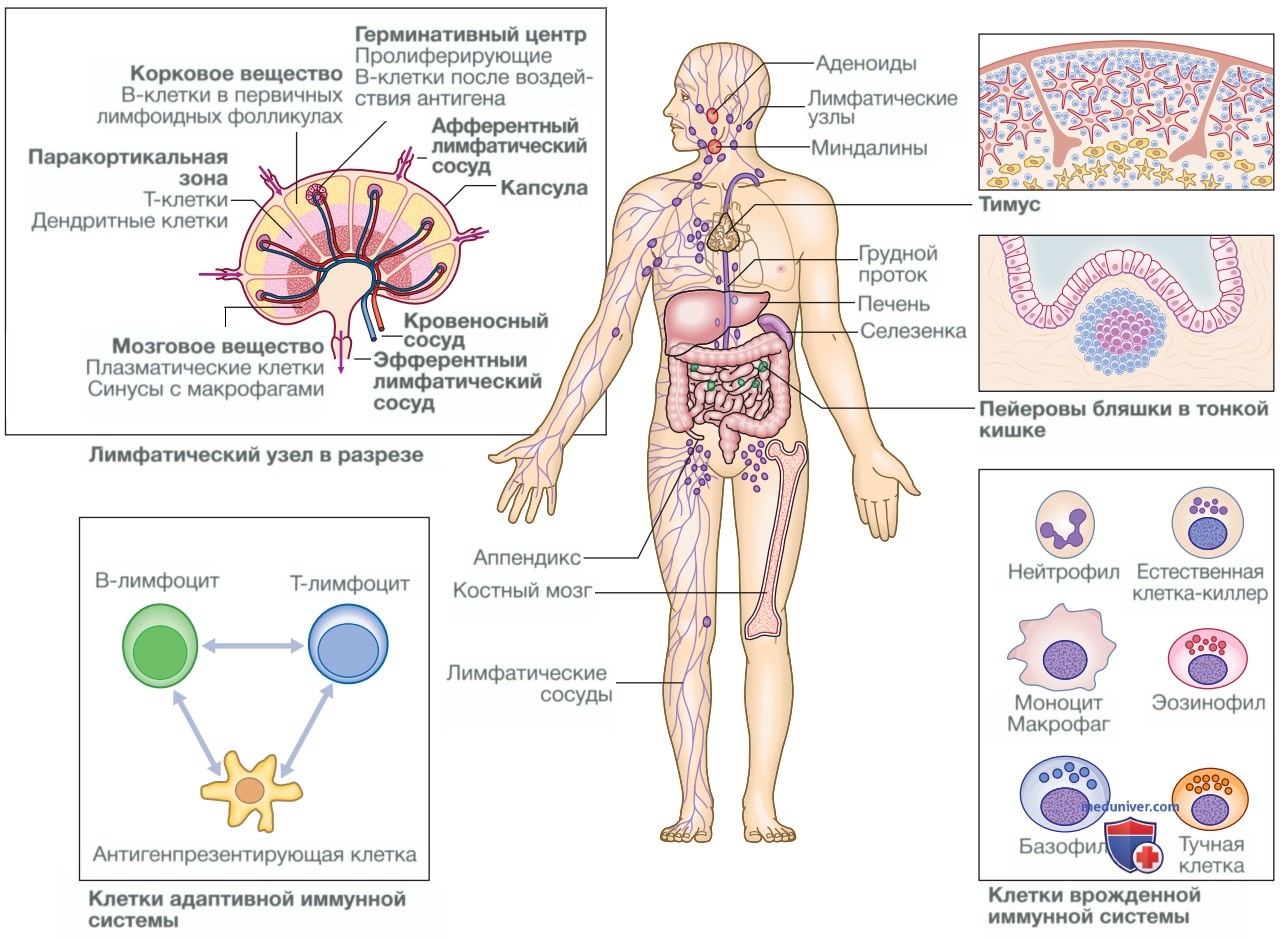
Иммунная система состоит из сложно взаимосвязанной сети лимфоидных органов, клеток и белков, имеющих четкое стратегическое расположение в целях защиты организма от инфекций (рис. 1). Иммунную защиту обычно подразделяют на врожденный иммунный ответ, который обеспечивает немедленную защиту от возбудителя, и адаптивный, или приобретенный, иммунный ответ, для формирования которого необходимо определенное время. Вместе с тем адаптивный, или приобретенный, иммунный ответ обеспечивает высокую специфичность и длительную защиту организма.
Рисунок 1. Анатомия иммунной системы
Врожденная защита от инфекции включает анатомические барьеры, фагоциты, растворимые молекулы, такие как белки комплемента и острой фазы, и естественные клетки-киллеры. Врожденная иммунная система распознает общие микробные структуры, присутствующие в тканях организмов, не относящихся к млекопитающим, и может быть мобилизована в течение нескольких минут. Специфический стимул будет вызывать практически идентичные ответы у разных индивидуумов, в отличие от адаптивных ответов антител и Т-клеток, которые у них существенно различаются.
а) Физические барьеры. Плотно соединенные кератиноциты кожи физически ограничивают колонизацию микроорганизмами. Гидрофобные жиры, выделяющиеся сальными железами, дополнительно отталкивают воду и микроорганизмы, а рост последних подавляется низким pH кожи и низким давлением кислорода. Пот также содержит лизоцим (фермент, который разрушает структурную целостность бактериальных клеточных стенок), аммиак, обладающий антибактериальными свойствами, и несколько противо-микробных пептидов, таких как дефензины. Точно так же слизистые оболочки дыхательной системы, желудочно-кишечного тракта и мочеполовых путей являются защитным физическим барьером для инфекции.
Секретируемая слизь захватывает попадающие на нее патогенные микроорганизмы, а иммуноглобулин (Ig) А, синтезируемый адаптивной иммунной системой, предотвращает прикрепление бактерий и вирусов к эпителиальным клеткам и их проникновение внутрь. Как и в коже, лизоцим и противомикробные пептиды в слизистых оболочках непосредственно уничтожают попадающих в организм возбудителей, а лактоферрин вызывает у бактерий голодание, связанное с отсутствием железа. В дыхательных путях реснички эпителия непосредственно задерживают патогенные микроорганизмы и способствуют их удалению со слизью, выделяющейся при чиханье и кашле.
В желудочно-кишечном тракте соляная кислота и амилаза слюны химически уничтожают бактерии, в то время как нормальная перистальтика и вызванная рвота или диарея помогают очищать организм от попавших в него микроорганизмов.
Микробном, состоящий из эндогенных комменсальных бактерий, обеспечивает дополнительную постоянную защиту от инфекций. Приблизительно 1014 бактерий обычно находятся на эпителиальных поверхностях в симбиозе с человеческим организмом. Они конкурируют с патогенными микроорганизмами за ограниченные ресурсы, включая пространство и питательные вещества, и образуют жирные кислоты и бактериолизины, которые ингибируют рост многих патогенных микроорганизмов. Кроме того, недавние исследования показали, что комменсальные бактерии помогают формировать иммунный ответ, индуцируя специфические регуляторные Т-клетки в кишечнике.
Эрадикация нормальной микрофлоры антибиотиками широкого спектра действия обычно приводит к развитию оппортунистической инфекции, вызванной такими микроорганизмами, как Clostridium difficile, которые быстро колонизируют незащищенную экологическую нишу.
Эти конститутивные барьеры очень эффективны, но, если внешняя защита нарушена появлением раневой поверхности или патогенным микроорганизмом, происходит активация специфических растворимых белков и клеток врожденной иммунной системы.
б) Фагоциты. Фагоциты («поедающие клетки») — это специализированные клетки, которые поглощают и убивают микроорганизмы, удаляют остатки клеток и инфекционных возбудителей и образуют воспалительные молекулы, которые регулируют другие компоненты иммунной системы. К ним относятся нейтрофилы, моноциты и макрофаги, и они особенно важны для защиты от бактериальных и грибковых инфекций. Фагоциты экспрессируют широкий спектр поверхностных рецепторов, включая рецепторы опознавания паттерна (pattern recognition receptors — PRR), которые распознают патоген-ассоциированные молекулярные паттерны (pathogen-associated molecular patterns — РАМР) на патогенных микроорганизмах, позволяя их идентифицировать.
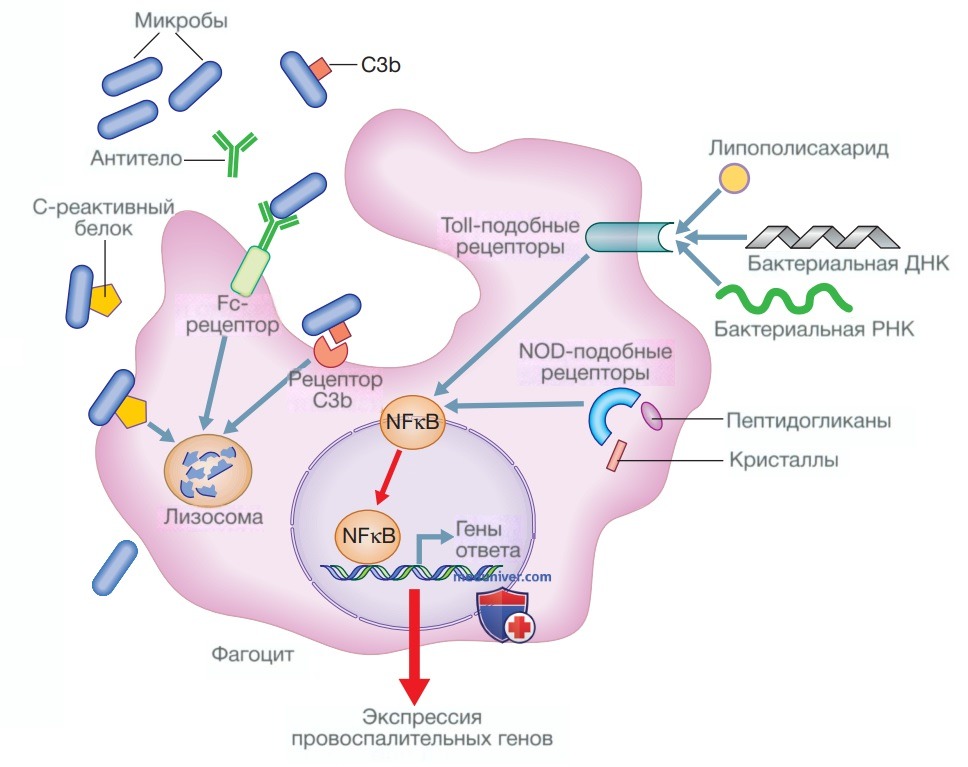
PPR включают Toll-подобные рецепторы, рецепторы, подобные доменам нуклеотидной олигомеризации (nucleotide oligomerization domaine — NOD), и рецепторы маннозы. РАМР, которые они распознают, представляют собой макромолекулы, отсутствующие в клетках млекопитающих, но свойственные одновременно целым группам микроорганизмов, включая компоненты бактериальной клеточной стенки, бактериальную дезоксирибонуклеиновую кислоту (ДНК) и вирусную двухцепочечную рибонуклеиновую кислоту (РНК). В то время как фагоциты могут распознавать микроорганизмы только через PPR, поглощение микроорганизмов значительно усиливается за счет опсонизации. Опсонины включают острофазовые белки, синтезируемые в печени, такие как С-реактивный белок (СРБ) и комплемент. Антитела, продуцирующиеся адаптивной иммунной системой, также действуют как опсонины. Они связываются как с патогеном, так и с рецепторами фагоцитов, выступая в качестве связующего звена между ними для облегчения фагоцитоза (рис. 2).
Рисунок 2. Фагоцитоз и опсонизация. Фагоцитоз микроорганизмов может усиливаться несколькими опсонинами, такими как С-реактивный белок, антитела и фрагменты комплемента, например СЗb, которые увеличивают способность фагоцитарных клеток поглощать микроорганизмы и уничтожать их. Фагоциты также распознают такие компоненты микроорганизмов, как липополисахарид, пептидогликаны, ДНК и РНК, в совокупности называемые патоген-ассоциированными молекулярными паттернами. Они активируют рецепторы опознавания паттерна, такие как Toll-подобные рецепторы и рецепторы, подобные доменам нуклеотидной олигомеризации, которые способствуют экспрессии воспалительных генов через путь ядерного фактора каппа-бета (Nuclear Factor kappa В — NFχB). Мочевая кислота и другие кристаллы также могут провоцировать воспаление через путь NOD
За этим следует внутриклеточное разрушение микроорганизма и последующая активация провоспалительных генов, что приводит к образованию провоспалительных цитокинов (см. ниже).
1. Нейтрофилы. Нейтрофилы, также называемые полиморфноядерными лейкоцитами, образуются в костном мозге и свободно циркулируют в крови. Они живут недолго (период полужизни — 6 ч) и образуются со скоростью 1011 клеток в сутки. Их функции заключаются в том, чтобы уничтожать микроорганизмы, способствовать быстрому прохождению клеток через ткани и неспецифически усиливать иммунный ответ. Эти функции опосредованы ферментами, содержащимися в гранулах, которые также обеспечивают внутриклеточную среду для уничтожения и разрушения микроорганизмов.
Различают два основных типа гранул: первичные, или азурофильные, и более многочисленные вторичные, или специфические. Первичные гранулы содержат миелопероксидазу и другие ферменты, играющие важную роль во внутриклеточном уничтожении и переваривании поглощенных микроорганизмов. Вторичные гранулы меньше по размеру и содержат лизоцим, коллагеназу и лактоферрин, которые могут высвобождаться во внеклеточное пространство. Синтез фермента увеличивается в ответ на инфекцию, что отражается в более интенсивном окрашивании гранул при микроскопии — это называется токсической зернистостью.
Изменения в поврежденных или инфицированных клетках запускают локальный синтез воспалительных молекул и цитокинов. Эти цитокины стимулируют образование и созревание нейтрофилов в костном мозге и их выход в кровоток. Нейтрофилы рекрутируются в определенные очаги инфекции под действием хемотаксических веществ, таких как интерлейкин-8 (интерлейкин — ИЛ), и за счет активации местного эндотелия. Увеличение экспрессии молекул клеточной адгезии на нейтрофилах и эндотелии также способствует миграции нейтрофилов. Выход нейтрофилов в кровоток приводит к увеличению их числа в периферической крови, что отмечается при ранней стадии инфекции. Оказавшись в инфицированной ткани, активированные нейтрофилы обнаруживают и поглощают попавшие в нее микроорганизмы. Первоначально микроорганизмы заключаются в мембраносвязанные везикулы, которые сливаются с цитоплазматическими гранулами с образованием фаголизосомы.
Внутри этого защищенного пространства происходит уничтожение микроорганизма за счет комбинации окислительного и неокислительного цитолиза. Окислительное уничтожение, также известное как кислородный взрыв, опосредовано ферментным комплексом никотинамидадениндинуклеотидфосфатоксидазы (НАДФН-оксидазы), который превращает кислород в активные и смертельные для микроорганизмов формы, такие как перекись водорода и супероксид. Фермент миелопероксидаза в нейтрофилах образует хлорноватистую кислоту, являющуюся мощным окислителем и противомикробным веществом. Неокислительное (не зависящее от кислорода) уничтожение происходит посредством высвобождения бактерицидных ферментов в фаголизосому. Каждый фермент имеет собственный противомикробный спектр, обеспечивая широкую активность против бактерий и грибков.
Дополнительной, недавно идентифицированной формой уничтожения микроорганизмов нейтрофилами является образование нейтрофильной внеклеточной ловушки. Активированные нейтрофилы могут высвобождать хроматин с гранулированными белками, такими как эластаза, с образованием внеклеточного матрикса, который связывается с микробными белками. Это может обездвижить или убить микроорганизмы без необходимости фагоцитоза. Процесс фагоцитоза и формирования нейтрофильной внеклеточной ловушки истощает запасы гликогена в нейтрофилах, что сопровождается их гибелью. Когда клетки погибают, их содержимое высвобождается, и лизосомальные ферменты разрушают коллаген и другие компоненты интерстиция, вызывая разжижение близлежащих тканей. Накопление погибших и отмирающих нейтрофилов приводит к образованию гноя, что при обширном процессе может закончиться образованием абсцесса.
2. Моноциты и макрофаги. Моноциты являются предшественниками тканевых макрофагов. Они образуются в костном мозге и попадают в кровоток, где составляют около 5% лейкоцитов. Из кровотока они мигрируют в периферические ткани, где дифференцируются в тканевые макрофаги и остаются в течение длительного периода времени. Специализированные популяции тканевых макрофагов включают клетки Купфера (Kupffer) в печени, альвеолярные макрофаги в легких, мезангиальные клетки в почках и клетки микроглии в головном мозге. Макрофаги, подобно нейтрофилам, способны к фагоцитозу и уничтожению микроорганизмов, но также играют важную роль в усилении и регуляции воспалительного ответа (табл. 1). Они особенно важны при тканевом надзоре и постоянно обследуют свое непосредственное окружение для обнаружения признаков повреждения тканей или вторжения микроорганизмов.
в) Дендритные клетки. Дендритные клетки представляют собой специализированные антигенпрезентирующие клетки, которые находятся в тканях, контактирующих с внешней средой, таких как кожа и слизистые оболочки. Они также могут обнаруживаться в незрелом состоянии в крови. Дендритные клетки исследуют свою окружающую среду для обнаружения посторонних частиц, а после активации переносят микробные антигены в регионарные лимфатические узлы, где они взаимодействуют с Т- и В-клетками для инициирования и формирования адаптивного иммунного ответа.
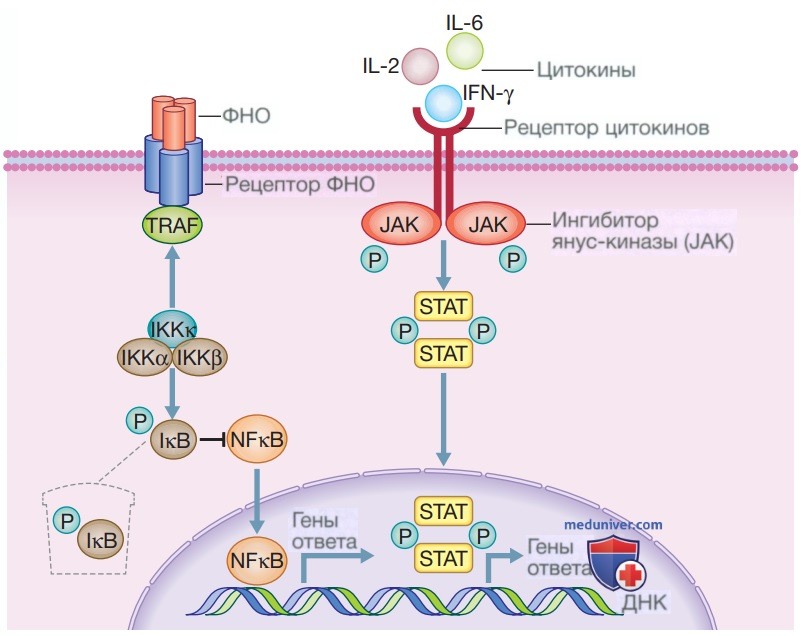
г) Цитокины. Цитокины являются сигнальными белками, которые синтезируются клетками иммунной системы и множеством других типов клеток. Идентифицировано более 100 цитокинов. Цитокины играют сложные и частично совпадающие роли в клеточном взаимодействии и регуляции иммунного ответа. Незначительные различия в выработке цитокинов, особенно при инициации иммунного ответа, могут оказать существенное влияние на исходы. Цитокины связываются со специфическими рецепторами на клетках-мишенях и активируют последующие внутриклеточные пути передачи сигналов, что в конечном итоге приводит к изменениям в транскрипции генов и функции клеток. Два важных пути передачи сигналов показаны на рис. 3. Путьядерного фактора каппа-бета (NFxB) активируется ФНО и другими членами суперсемейства ФНО, такими как активатор рецептора лиганда NFxB (RANK.L), Toll- и NOD-подобные рецепторы (см. рис. 2).
Рисунок 3. Пути передачи сигналов цитокинов и иммунный ответ. Цитокины регулируют иммунный ответ посредством связывания со специфическими рецепторами, которые активируют множество внутриклеточных путей передачи сигналов, два из которых показаны на рисунке. Члены суперсемейства фактора некроза опухоли, Toll- и NOD-подобные рецепторы (см. рис. 2) передают сигналы через путь ядерного фактора каппа В. Некоторые другие цитокины, в том числе ИЛ-2, ИЛ-6 и интерфероны, используют путь янус-киназы/переносчиков сигнала и активатора транскрипции [для регуляции функции клеток (см. текст для получения более подробной информации)]. IχB — ингибитор каппа В; IKK — ингибитор каппа В киназы; Р — фосфорилирование сигнального белка; TRAF — фактор, ассоциированный с рецептором фактора некроза опухоли
Что касается членов суперсемейства ФНО, связывание рецептора приводит к тому, что ингибитор комплекса каппа В киназы (Inhibitor of Карра В Kinase complex — IKK), состоящий из трех белков, рекрутируется в рецептор за счет связывания белков, ассоциированных с рецептором ФНО (Tumor necrosis factor Receptor-Associated proteins Factor — TRAF). Это активирует IKK, что в свою очередь приводит к фосфорилированию ингибитора белка ядерного фактора каппа В (Inhibitor of nuclear factor kappa Bprotein — IxB), вызывая его деградацию и позволяя NFxB транслоцироваться в ядро и активировать транскрипцию генов. Путь янус-киназы/ переносчиков сигнала и активатора транскрипции (Janus kinase/Signal Transducers and Activators of Transcriptions — JAK-STAT) участвует в передаче сигналов со многих рецепторов цитокинов, включая сигналы для ИЛ-2, ИЛ-6 и γ-интерферона(γ-ИФН).
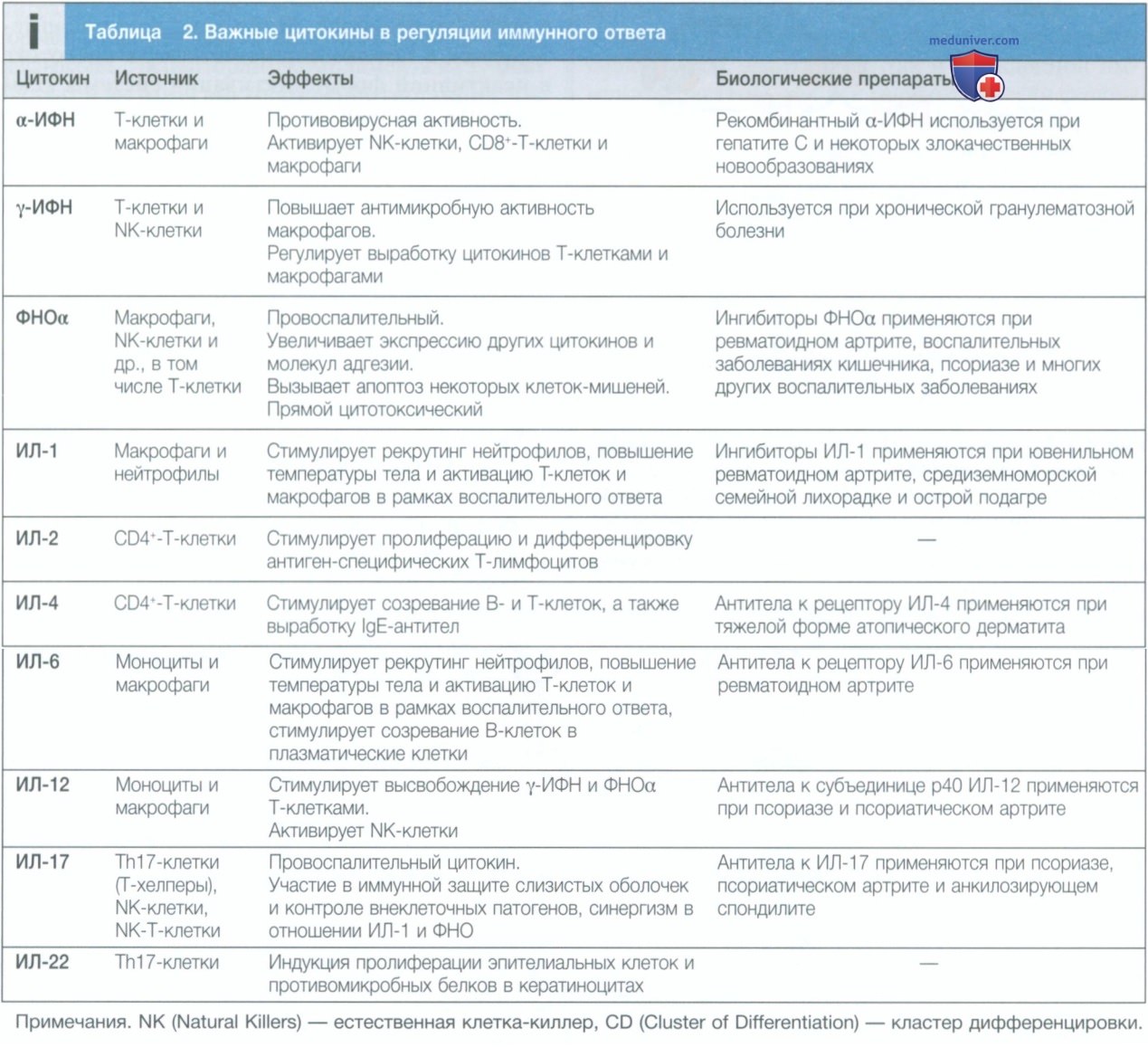
При связывании рецептора белки янус-киназы (Janus kinase — JAK рекрутируются во внутриклеточную часть рецептора и фосфорилируются. В свою очередь, они фосфорилируют белки STAT), которые транслоцируются в ядро и активируют транскрипцию генов, изменяя функцию клеток. Функция и взаимосвязи некоторых важных цитокинов с заболеваниями перечислены в табл. 2. Ингибиторы цитокинов в настоящее время рутинно используются при лечении аутоиммунных заболеваний, большинство из них представляют собой моноклональные антитела к цитокинам или их рецепторам. Кроме того, были разработаны низкомолекулярные ингибиторы, которые блокируют внутриклеточные пути передачи сигналов, используемые цитокинами. К ним относятся ингибиторы янус-киназы тофацитиниб и барацитиниб, используемые при лечении ревматоидного артрита, и ингибитор тирозинкиназы иматиниб, применяемый при хроническом миелолейкозе.
д) Интегрины. Интегрины представляют собой трансмембранные белки, которые играют важную роль во взаимодействиях клеток друг с другом и с матриксом. Они опосредуют прикрепление клетки к внеклеточному матриксу, передачу сигнала и миграцию клеток. Их роль в аутоиммунных заболеваниях была подробно изучена. Таргетная терапия с использованием рекомбинантного гуманизированного антитела к α4-интегрину — натализумаба является эффективным средством для лечения рассеянного склероза, которое предотвращает проникновение иммунных клеток в эндотелий сосудов и центральную нервную систему (ЦНС).
е) Комплемент. Система комплемента включает группу, состоящую более чем из 20 строго регулируемых, функционально связанных белков, которые стимулируют воспаление и элиминируют попавшие в организм патогены. Белки комплемента синтезируются в печени и присутствуют в кровотоке в неактивной форме. Активация системы комплемента приводит к быстрому развертыванию биологического каскада, аналогичного каскаду свертывания крови.
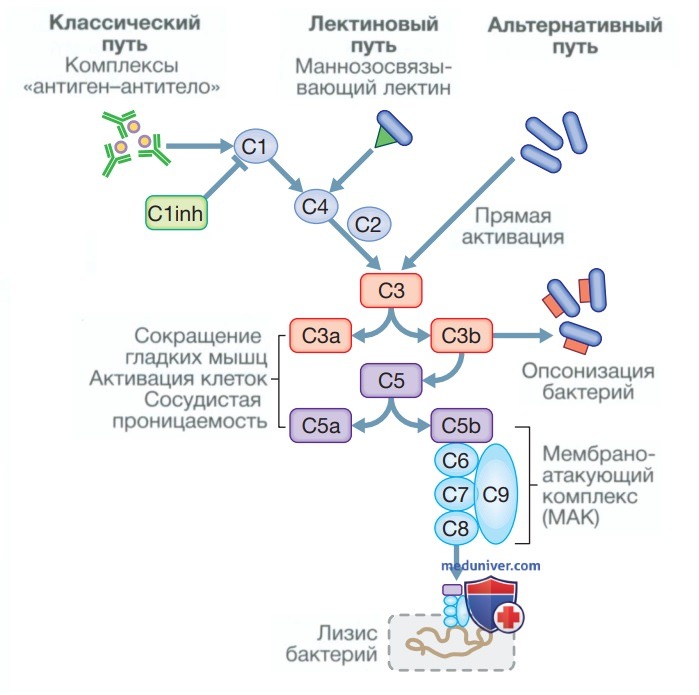
Выделяют три механизма, способные активировать каскад комплемента (рис. 4).
Рисунок 4. Путь комплемента. Классический путь активируется связыванием комплексов «антиген-антитело» с С1, а блокируется ингибитором С1 (С1 inhibitor — С1 inh), тогда как маннозосвязывающие лектины, представляющие собой макромолекулы, связывающиеся с различными микроорганизмами, активируют этот путь при связывании С4. Бактерии могут напрямую активировать путь через СЗ, который играет ключевую роль в активации комплемента всеми тремя путями
• Альтернативный путь запускается непосредственно связыванием С3 с компонентами бактериальной клеточной стенки, такими как липополисахарид грамотрицательных бактерий и тейхоевая кислота грамположительных бактерий.
• Классический путь запускается, когда две молекулы или более IgM- или IgG-антител связываются с антигеном. Вызванное этим процессом конформационное изменение обнажает участки связывания на антителах к первому белку в классическом пути — С1, который представляет собой разветвленную молекулу, способную связывать до шести молекул антител. Как только две «головы» молекулы С1 или более связываются с антителом, запускается классический каскад. Важным ингибитором классического пути является ингибитор Cl (Clinh), как показано на рис. 4.
• Лектиновый путь активируется непосредственным связыванием маннозосвязывающего лектина с углеводами микробной клеточной поверхности. Это имитирует связывание С1 с иммунными комплексами и напрямую стимулирует классический путь, минуя необходимость формирования иммунных комплексов.
Активация комплемента любым из этих путей приводит к активации С3. Это, в свою очередь, активирует последний общий путь, в котором белки комплемента С5—С9 собираются в единую структуру, образуя мембраноатакующий комплекс. Он способен проколоть клеточную стенку, что приводит к осмотическому лизису клеток-мишеней. Данный этап особенно важен для защиты от инкапсулированных бактерий, таких как Neisseria spp. и Haemophilus influenzae.
Фрагменты комплемента, образующиеся при активации каскада, также могут действовать как опсонины, повышая восприимчивость микроорганизмов к фагоцитозу макрофагами и нейтрофилами (см. рис. 2). Кроме того, они являются хемотаксическими веществами, способствующими миграции лейкоцитов в места воспаления. Некоторые фрагменты действуют как анафилотоксины, связываясь с рецепторами комплемента на тучных клетках и вызывая высвобождение гистамина, что увеличивает проницаемость сосудов. Продукты активации комплемента также помогают направлять иммунные комплексы на антигенпрезентирующие клетки, обеспечивая связь между врожденной и приобретенной иммунной системами. Наконец, активированные продукты комплемента растворяют иммунные комплексы, которые запускают каскад, минимизируя повреждение окружающих тканей.
Моноклональное антитело, направленное против центральной молекулы комплемента С5, — экулизумаб — был разработан для терапевтического применения при пароксизмальной ночной гемоглобинурии и атипичных гемолитико-уремических синдромах. Сообщалось об инфекционных осложнениях, включая менингококковый сепсис, на фоне терапии экулизумабом, что подчеркивает важность системы комплемента в профилактике таких инфекций.
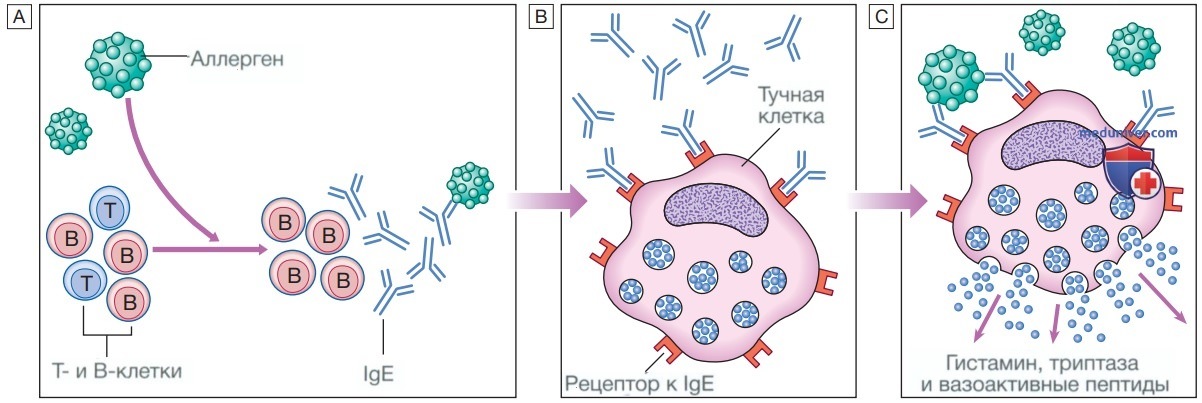
е) Тучные клетки и базофилы. Тучные клетки и базофилы образуются в костном мозге и играют центральную роль в аллергических реакциях. Тучные клетки находятся преимущественно в тканях, подвергающихся воздействию внешней среды, таких как кожа и кишечник, в то время как базофилы циркулируют в периферической крови и рекрутируются в ткани в ответ на воспаление. Тучные клетки и базофилы содержат большие цитоплазматические гранулы, в которых находятся вазоактивные вещества, такие как гистамин (см. рис. ниже). Тучные клетки и базофилы экспрессируют рецепторы к IgE на своей клеточной поверхности, с помощью которых происходит связывание IgE. При столкновении со специфическим антигеном клетка активируется и высвобождает гистамин и другие медиаторы, находящиеся в гранулах, а также начинает синтезировать дополнительные медиаторы, включая лейкотриены, простагландины и цитокины. Инициируется воспалительный каскад, который усиливает местный кровоток и проницаемость сосудов, стимулирует сокращение гладких мышц и увеличивает секрецию на поверхности слизистых оболочек.
Реакция гиперчувствительности 1-го типа (немедленного): А — после контакта с аллергеном В-клетки продуцируют к нему антитела класса Е (IgE); Б — специфические IgE связываются с высокоаффинными рецепторами на поверхности циркулирующих тучных клеток: В — при повторном контакте аллерген связывается с IgE, покрывающими тучные клетки. Это перекрестная сшивка IgE-антигеном запускает активацию тучных клеток с высвобождением вазоактивных медиаторов
ж) Естественные клетки-киллеры. NK-клетки представляют собой большие гранулярные лимфоциты, которые играют важную роль в защите от опухолей и вирусов. Они проявляют свойства как адаптивной, так и врожденной иммунной системы, так как морфологически сходны с лимфоцитами и распознают сходные лиганды, однако они не являются антиген-специфически-ми и не могут формировать иммунологическую память. NK-клетки экспрессируют множество рецепторов на клеточной поверхности, часть из которых являются стимулирующими, а другие — ингибирующими. В норме эффекты ингибирующих рецепторов преобладают. Они распознают молекулы человеческого лейкоцитарного антигена (Human Leucocyte Antigen — HLA), которые экспрессируются на нормальных ядросодержащих клетках, предотвращая опосредованную NK-клетками атаку, тогда как стимулирующие рецепторы распознают молекулы, экспрессирующиеся, главным образом, при повреждении клеток. Это позволяет NK-клеткам оставаться толерантными к здоровым клеткам, но не к поврежденным.
Когда клетки инфицированы вирусами или подвергаются злокачественным изменениям, экспрессия молекул HLA I класса на клеточной поверхности может уменьшаться. Это важный механизм, с помощью которого данные клетки избегают адаптивных ответов Т-лимфоцитов. Однако в таких условиях защитное действие NK-клеток становится важным, поскольку подавление HLA I класса аннулирует ингибирующие сигналы, которые обычно предотвращают активацию NK-клеток. Конечным результатом является атака NK-клеток на патологическую клетку-мишень. NK-клетки также могут активироваться при связывании комплексов «антиген—антитело» с поверхностными рецепторами. Образуется физическая связь NK-клетки с ее мишенью аналогично процессу опсонизации, и этот процесс носит название антителозависимой клеточной цитотоксичности.
Активированные NK-клетки могут уничтожать свои мишени различными способами. Они секретируют порообразующие белки, такие как перфорин, в мембрану клетки-мишени и протеолитические ферменты, называемые гранзимами, в саму клетку-мишень, что вызывает апоптоз. Кроме того, NK-клетки образуют различные цитокины, такие как ФНОа и γ-ИФН, которые оказывают прямое антивирусное и противоопухолевое действие.