Адаптивная иммунная система - кратко с точки зрения внутренних болезней
Если врожденная иммунная система не в состоянии обеспечить эффективную защиту от вторжения микроорганизма, мобилизуется адаптивная иммунная система (см. рис. 1), которая имеет три основные характеристики.
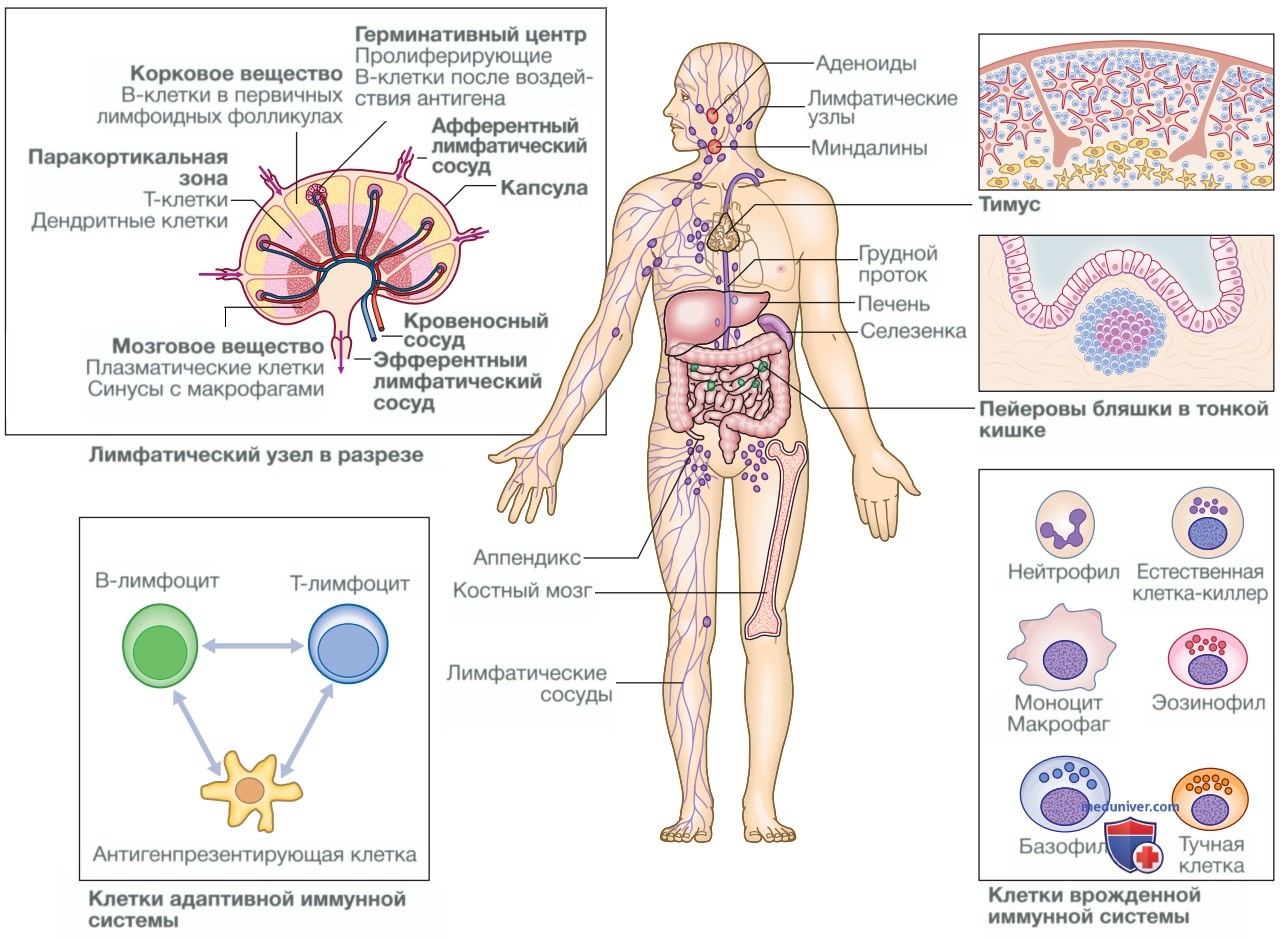
Рисунок 1. Анатомия иммунной системы
• Обладает исключительной специфичностью и может дифференцировать очень небольшие различия в молекулярной структуре.
• Высокоадаптивна и может реагировать практически на неограниченное количество молекул.
• Обладает иммунологической памятью, а изменения, вызванные первоначальной активацией антигеном, позволяют вырабатывать более эффективный иммунный ответ при последующих контактах с ним.
Выделяют два основных направления адаптивного иммунного ответа. Гуморальный иммунитет включает выработку антител В-лимфоцитами, а клеточный иммунитет подразумевает активацию Т-лимфоцитов, синтезирующих и высвобождающих цитокины, которые влияют на другие клетки, а также непосредственно уничтожают клетки-мишени. Эти две ветви тесно взаимодействуют друг с другом и с компонентами врожденной иммунной системы, чтобы достичь максимальной эффективности иммунного ответа.
а) Лимфоидные органы. Первичные лимфоидные органы участвуют в развитии лимфоцитов. Они включают костный мозг, в котором Т- и В-лимфоциты дифференцируются из гемопоэтических стволовых клеток и где также созревают В-лимфоциты, и тимус, являющийся местом созревания Т-клеток (см. рис. 1). После созревания лимфоциты мигрируют во вторичные лимфоидные органы.
К ним относятся селезенка, лимфатические узлы и лимфоидная ткань слизистых оболочек. Они захватывают и концентрируют чужеродные вещества и являются основными участками взаимодействия интактных лимфоцитов и микроорганизмов.
1. Тимус. Тимус представляет собой двудольную структуру, располагающуюся в переднем средостении, и состоит из коркового и мозгового вещества. Кора густо заселена незрелыми Т-клетками, которые мигрируют в мозговое вещество, чтобы пройти отбор и созревание. Тимус наиболее активен в период внутриутробного развития и у новорожденных, а после полового созревания подвергается инволюции.
Врожденное отсутствие тимуса сопровождается тяжелым Т-клеточным иммунодефицитом, но хирургическое удаление тимуса в детском возрасте (обычно во время обширных кардиологических операций) не приводит к серьезной дисфункции иммунной системы.
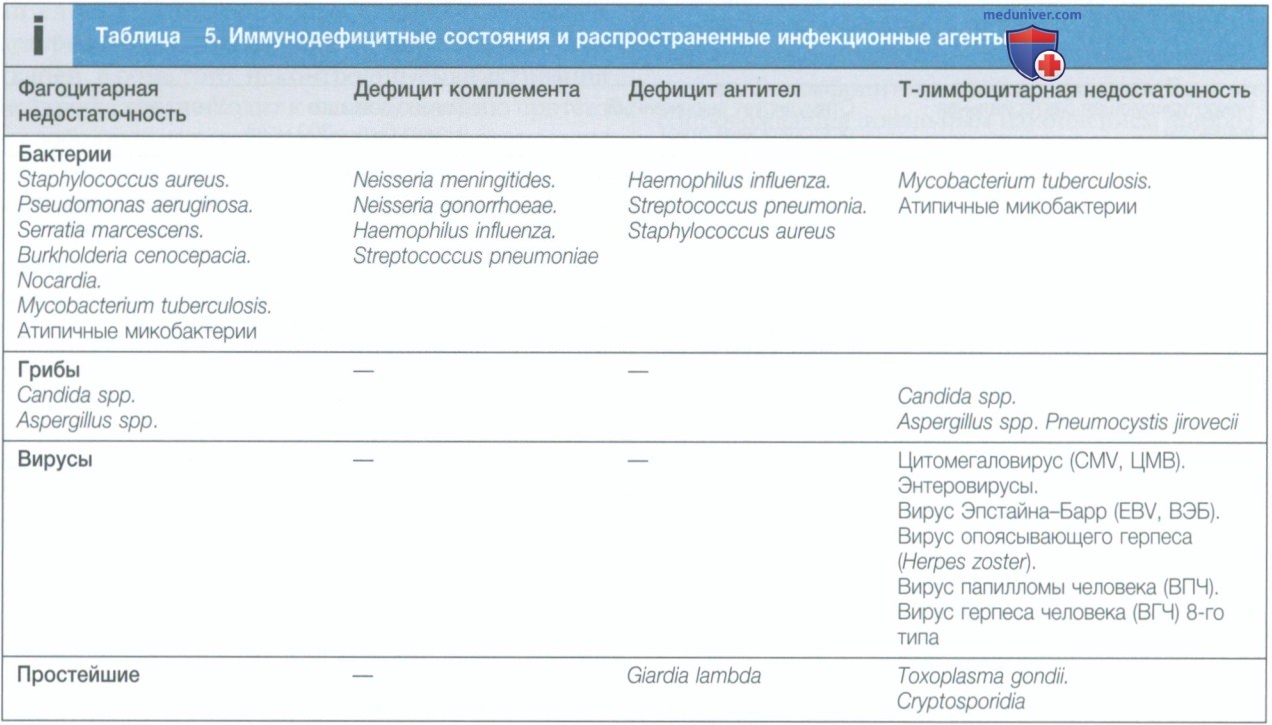
2. Селезенка. Селезенка — крупнейший из вторичных лимфоидных органов. Она очень эффективна при фильтрации крови и является важной областью фагоцитоза стареющих эритроцитов, бактерий, иммунных комплексов и других продуктов жизнедеятельности клеток, а также местом синтеза антител. Селезенка важна для защиты от инкапсулированных бактерий, и у лиц с аспленией повышен риск генерализованной инфекции, вызванной Streptococcus pneumoniae и Н. influenzae (см. табл. 5).
3. Лимфатические узлы. Лимфатические узлы расположены таким образом, чтобы получать максимальный отток лимфы из областей внешнего контакта, и довольно сложно устроены (см. рис. 1).
• Кора содержит первичные лимфоидные фолликулы, которые являются местом взаимодействия В-лимфоцитов. Когда В-клетки сталкиваются с антигеном, они подвергаются интенсивной пролиферации, образуя герминативные центры.
• Паракортикальная зона богата Т-лимфоцитами и дендритными клетками.
• Мозговое вещество — основное место нахождения плазматических клеток, секретирующих антитела.
• В мозговом веществе находится много синусов, содержащих большое количество макрофагов.
4. Лимфоидная ткань слизистых оболочек. Лимфоидная ткань слизистых оболочек (Mucosa-Associated Lymphoid Tissue — MALT) состоит из диффузно распределенных вдоль поверхности слизистой оболочки лимфоидных клеток и фолликулов. Ее функция аналогична функциям более организованных инкапсулированных лимфатических узлов. Это миндалины, аденоиды и пейеровы бляшки в тонкой кишке.
5. Лимфатические сосуды. Лимфоидная ткань связана сетью лимфатических сосудов, имеющих три основные функции: они обеспечивают доступ к лимфатическим узлам, возвращают интерстициальную жидкость в венозную систему и транспортируют жир из тонкой кишки в системный кровоток (см. рис. ниже). Лимфатические сосуды начинаются как слепые капилляры, которые, сливаясь, образуют лимфатические протоки, входящие и выходящие из регионарных лимфатических узлов в виде афферентных и эфферентных протоков соответственно.
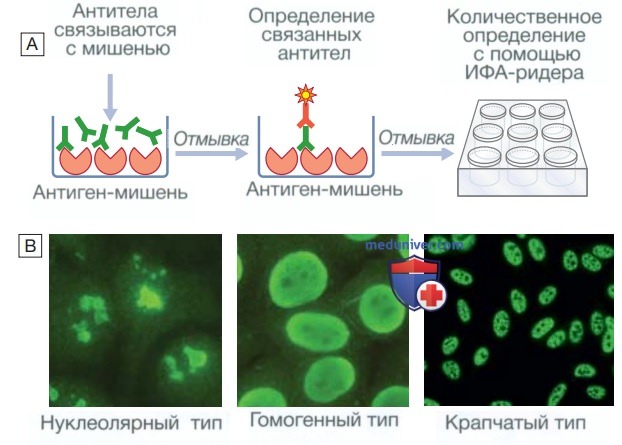
Исследование на аутоантитела: А — определение уровня антител с помощью иммуносорбентного анализа с ферментной меткой. На планшеты для микротитрования адсорбируется исследуемый антиген, а затем добавляется сыворотка крови пациента. При наличии в образце сыворотки аутоантитела связываются с антигеном-мишенью, фиксированным на планшете. Количество связанных антител определяют с помощью добавления детектирующих антител, связанных с ферментом-меткой, который окрашивает бесцветный субстрат, что выявляется с помощью инфракрасного ридера; В — качественный анализ аутоантител на основании типа ядерного свечения. При данном исследовании к культивированным клеткам добавляют сыворотку крови пациента и детектирующие антитела с флуоресцентной меткой, что позволяет выявить связавшиеся с антигеном антиядерные антитела, которые обнаруживаются в виде ярко-зеленого свечения. При различных заболеваниях соединительной ткани обнаруживают разные типы иммунофлуоресцентного свечения.
В конечном итоге они объединяются и впадают в грудной проток и левую подключичную вену. Лимфатические узлы могут быть глубокими и поверхностными, располагаясь по ходу основных кровеносных сосудов.
б) Гуморальный иммунитет. Гуморальный иммунитет опосредован В-лимфоцитами, которые дифференцируются из гемопоэтических стволовых клеток в костном мозге. Их основные функции — вырабатывать антитела и взаимодействовать с Т-клетками, но они также участвуют в презентации антигена. Зрелые В-лимфоциты могут находиться в костном мозге, лимфоидной ткани, селезенке и гораздо реже — в кровотоке. Они экспрессируют уникальный иммуноглобулиновый рецептор на своей клеточной поверхности, В-клеточный рецептор, который связывается с растворимыми антигенными мишенями (рис. 5).
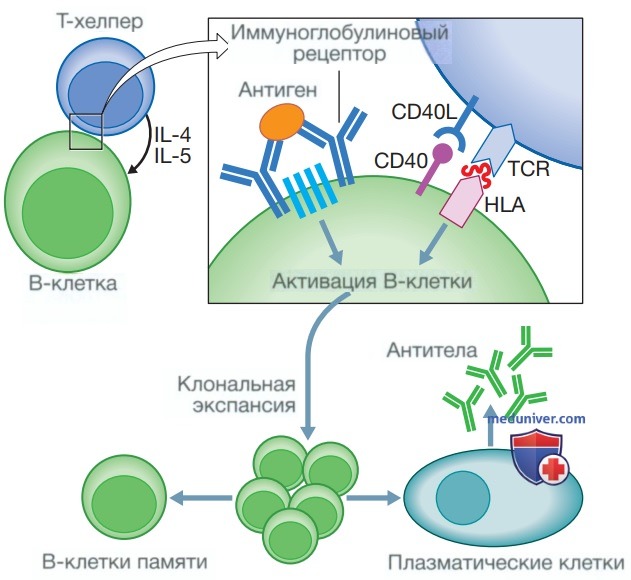
Рисунок 5. Активация В-клеток. Активация В-клеток инициируется связыванием антигена с иммуноглобулиновым рецептором на поверхности клетки. Для продолжения активации также необходимо взаимодействие с Т-хелперами, обеспечивающими дополнительные сигналы посредством связывания лиганда CD40 (CD40L) с его рецептором CD40; взаимодействие между Т-клеточным рецептором (T-Cell Receptor — TCR) и антигенными пептидами, представленными молекулами HLA на поверхности В-клетки, и цитокинами, высвобождаемыми Т-хелперами. Полностью активированные В-клетки подвергаются клональной экспансии с дифференцировкой в плазматические клетки, которые образуют антитела. После активации создаются клетки памяти, позволяющие быстро образовывать антитела, когда тот же самый антиген встречается повторно. CD — кластер дифференцировки; ИЛ — интерлейкин
Контакт с антигеном обычно происходит в лимфатических узлах. При наличии соответствующих цитокинов и других сигналов от близлежащих Т-лимфоцитов антиген-специфические В-клетки реагируют быстрой пролиферацией с развитием процесса, известного как клональная экспансия (см. рис. 5). Это сопровождается очень сложной серией генетических перестроек, которая носит название соматической сверхмутации, в ходе чего возникает популяция В-клеток, экспрессирующих рецепторы с большей аффинностью к антигену, чем у исходной популяции. Эти клетки дифференцируются либо в долгоживущие клетки памяти, которые находятся в лимфатических узлах, либо в плазматические клетки, синтезирующие антитела.
Клетки памяти позволяют осуществлять более быстрый и эффективный ответ при последующем воздействии данного патогена.
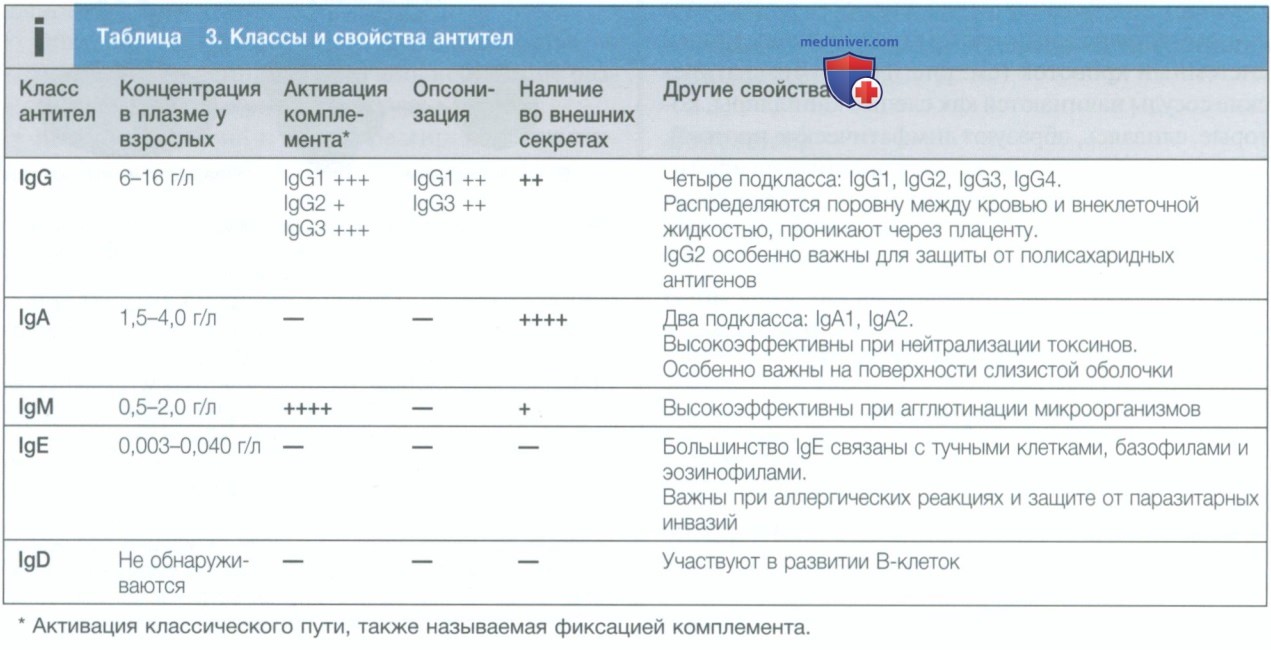
в) Иммуноглобулины. Иммуноглобулины играют центральную роль в гуморальном иммунитете. Они представляют собой растворимые белки, синтезируемые плазматическими клетками, и состоят из двух тяжелых и двух легких цепей (рис. 6). Тяжелая цепь определяет класс, или изотип, антитела, например IgG, IgA, IgM, IgE или IgD. Также выделяют подклассы IgG и IgA. Антиген распознается антигенсвязывающими областями (Fab) как тяжелой, так и легкой цепи, а последствия связывания антител определяются константной областью тяжелой цепи (Fc) (табл. 3). Антитела имеют несколько функций.
Рисунок 6. Структура молекулы иммуноглобулина (антитела). Вариабельная область отвечает за связывание антигена, тогда как константная область может взаимодействовать с иммуноглобулиновыми рецепторами, экспрессируемыми на иммунных клетках
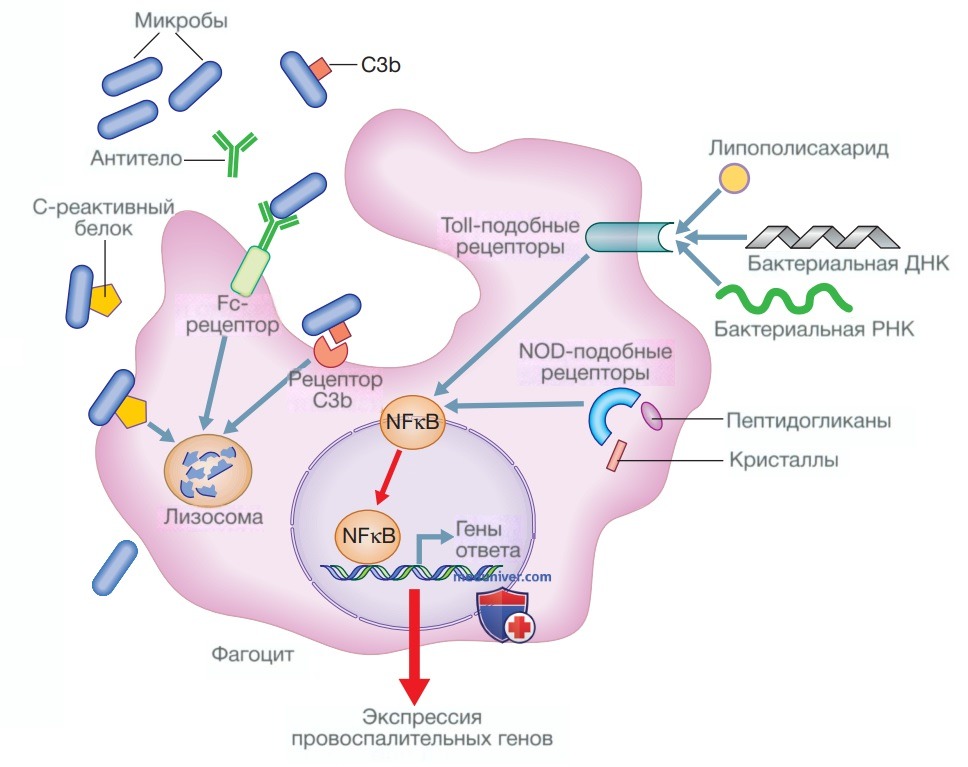
Они облегчают фагоцитоз, действуя как опсонины (см. рис. 2), и способствуют уничтожению клеток цитотоксическими клетками, особенно NK-клетками, посредством антителозависимой клеточной цитотоксичности. Связывание антител с антигеном может инициировать активацию классического пути комплемента (см. рис. 4). Кроме того, антитела способны непосредственно нейтрализовать биологическую активность их антигена-мишени. Это особенно важное свойство IgA-антител, которые действуют преимущественно на поверхности слизистой оболочки.
Рисунок 2. Фагоцитоз и опсонизация. Фагоцитоз микроорганизмов может усиливаться несколькими опсонинами, такими как С-реактивный белок, антитела и фрагменты комплемента, например СЗb, которые увеличивают способность фагоцитарных клеток поглощать микроорганизмы и уничтожать их. Фагоциты также распознают такие компоненты микроорганизмов, как липополисахарид, пептидогликаны, ДНК и РНК, в совокупности называемые патоген-ассоциированными молекулярными паттернами. Они активируют рецепторы опознавания паттерна, такие как Toll-подобные рецепторы и рецепторы, подобные доменам нуклеотидной олигомеризации, которые способствуют экспрессии воспалительных генов через путь ядерного фактора каппа-бета (Nuclear Factor kappa В — NFχB). Мочевая кислота и другие кристаллы также могут провоцировать воспаление через путь NOD
Гуморальный иммунный ответ характеризуется иммунологической памятью, при которой образование антител в ответ на последовательные воздействия антигена качественно и количественно улучшается по сравнению с первичным контактом. Когда ранее не стимулированный, или интактный, В-лимфоцит активируется антигеном, среди всех антител сначала образуются IgM, которые появляются в сыворотке крови через 5—10 дней. В зависимости от дополнительных стимулов, обеспечиваемых Т-лимфоцитами, другие классы антител (IgG, IgA и IgE) вырабатываются через 1—2 нед.
Если впоследствии В-клетка памяти повторно подвергается воздействию того же антигена, латентный период между контактом с антигеном и образованием антител сокращается до 2—3 дней, количество синтезируемых антител увеличивается, и в ответе преобладают IgG-антитела с высокой аффинностью. Кроме того, в отличие от первичного образования антител, вторичные ответы не требуют дополнительного участия Т-лимфоцитов. Это позволяет быстро генерировать высокоспецифичные ответы при повторном воздействии патогена, что является важным механизмом эффективной вакцинации.
г) Клеточный иммунитет. Клеточный иммунитет опосредован Т-лимфоцитами, которые имеют важное значение в защите от вирусов, грибов и внутриклеточных бактерий. Они также играют важную иммунорегуляторную роль, организуя и регулируя ответы других компонентов иммунной системы. Предшественники Т-лимфоцитов дифференцируются из гемопоэтических стволовых клеток в костном мозге и в незрелом состоянии экспортируются в тимус (см. рис. 1).
Отдельные Т-клетки экспрессируют уникальный рецептор, который является высокоспецифичным для одного антигена. Внутри тимуса Т-клетки проходит строгий отбор с разрушением аутореактивных клеток. Зрелые Т-лимфоциты покидают тимус и пролиферируют, заселяя другие органы иммунной системы. Было подсчитано, что у человека имеется 107—109 клонов Т-клеток, каждый из которых обладает уникальным Т-клеточным рецептором, обеспечивая по крайней мере частичное покрытие для любого встречающегося антигена.
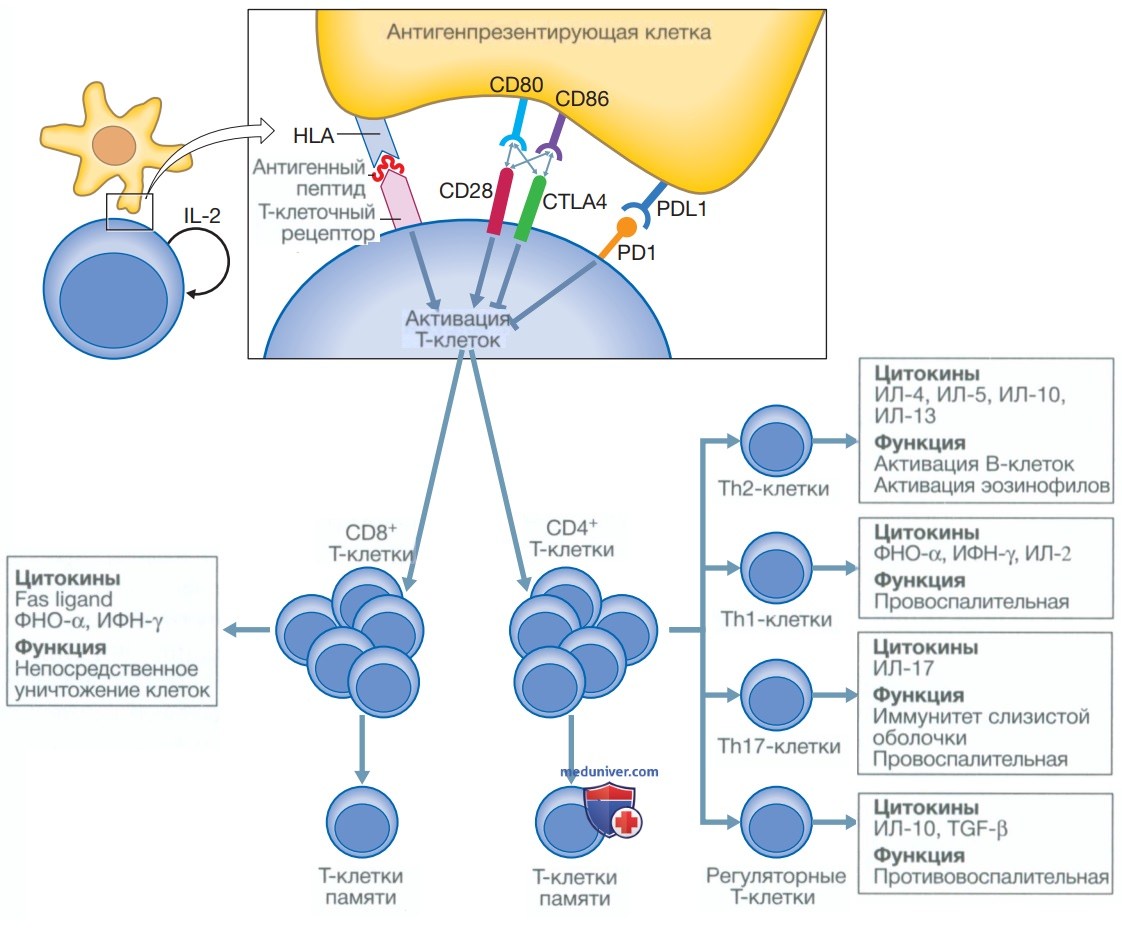
В отличие от В-клеток, Т-клетки не могут распознавать интактные белковые антигены в своей нативной форме. Вместо этого белок должен быть расщеплен на составляющие пептиды антигенпрезентирующими клетками для представления Т-лимфоцитам в связи с молекулами HLA на поверхности антигенпрезентирующих клеток. Это называется фрагментацией и презентацией антигена, и именно комплекс пептида и HLA в совокупности распознается отдельными Т-клетками (рис. 7). Структура молекул HLA значительно отличается у разных людей.
Рисунок 7. Активация Т-клеток. Активация Т-клеток инициируется, когда антигенный пептид, связанный с молекулой HLA на антигенпрезентирующих клетках, взаимодействует с Т-клеточным рецептором, экспрессируемым Т-лимфоцитами. Однако для активации Т-клеток требуются дополнительные сигналы. Они включают связывание костимулирующих молекул CD80 и CD86 с CD28 на Т-клетке и интерлейкином-2, который аутокринно вырабатывается активированными Т-клетками. Тем не менее есть и другие молекулы, которые могут ингибировать активацию Т-клеток, в том числе белок 4, ассоциированный с цитотоксическими Т-лимфоцитами {Cytotoxic T-Lymphocyte-Associated protein 4 — CTLA4), который конкурирует с CD28 за связывание с CD80 и CD86; и PD1 {Programmed cell Death 1) — белок запрограммированной гибели клеток 1, который, связываясь с PDL1 (Programmed Death Ligand 1) — лигандом белка запрограммированной гибели клеток 1, также оказывает ингибирующее действие. После активации Т-клетки пролиферируют и в зависимости от их подтипа выполняют различные функции с четкими паттернами образования цитокинов, как показано на рисунке. Также образуются клетки памяти, которые могут вызвать быстрый иммунный ответ при встрече с тем же антигеном. CD — кластер дифференцировки; CD40L — лиганд CD40; γ-ИФН — γ-интерферон; ИЛ — интерлейкин; TGFβ (Transforming Growth Factor β) — трансформирующий фактор роста β; ФНОα — фактор некроза опухоли α
Поскольку каждая молекула HLA обладает способностью представлять Т-лимфоцитам слегка отличающийся пептидный спектр, это обеспечивает огромное разнообразие в распознавании антигенов популяцией Т-клеток. Все ядросодержащие клетки обладают способностью фрагментировать и представлять антигены, но дендритные клетки, макрофаги и В-лимфоциты имеют специализированные антигенпрезентирующие функции. Они экспрессируют дополнительные костимулирующие молекулы, такие как CD80 и CD86, обеспечивая необходимый повторный сигнал для полной активации Т-клеток.
Т-лимфоциты можно разделить на две подгруппы на основании их функции и распознавания молекул HLA. Их обозначают как CD4+- и CD8+-Т-клетки в зависимости от количества антигенов кластера дифференцировки (Cluster of Differentiation — CD) основных белков, экспрессируемых на их клеточной поверхности.
1. CD8+-T-лимфоциты. Эти клетки распознают антигенные пептиды в связи с молекулами HLA I класса (HLA-A, HLA-B, HLA-С). Они напрямую уничтожают инфицированные клетки с помощью синтеза порообразующих молекул, таких как перфорин, и высвобождения переваривающих ферментов, запускающих апоптоз клетки-мишени, и особенно важны для защиты от вирусной инфекции.
2. CD4+-Т-лимфоциты. Эти клетки распознают пептиды, представленные на молекулах HLA II класса (HLA-DR, HLA-DP и HLA-DQ), и выполняют преимущественно иммунорегуляторные функции. Они синтезируют цитокины и обеспечивают костимулирующие сигналы, которые поддерживают активацию CD8+-T-лимфоцитов и помогают выработке зрелых антител В-клетками. Кроме того, их тесное взаимодействие с фагоцитами определяет продукцию цитокинов обоими типами клеток. Лимфоциты CD4+ могут быть дополнительно подразделены на виды в зависимости от синтезируемых ими цитокинов.
• Th1-клетки (Т-хелперы) обычно образуют ИЛ-2, γ-ИФН и ФНОα и поддерживают развитие реакций гиперчувствительности замедленного типа.
• Th2-клетки обычно синтезируют ИЛ-4, ИЛ-5, ИЛ-10 и ИЛ-13 и способствуют развитию аллергических реакций.
• Т-регуляторные клетки (T-reg) представляют собой дополнительную подгруппу специализированных CD4+-лимфоцитов, которые важны для активного подавления активации других клеток и предотвращения развития аутоиммунных заболеваний.
• Th17-клетки — провоспалительные клетки, определяемые по синтезу ими ИЛ-17. Они связаны с регуляторными Т-клетками и играют роль в иммунной защите на поверхности слизистых оболочек.
Активация Т-клеток регулируется балансом между костимулирующими молекулами, повторным сигналом, необходимым для активации, и ингибирующими молекулами, которые подавляют активность Т-клеток. Одна такая ингибирующая молекула, CTLA4, используется с терапевтической целью в виде препарата «Абатацепт», который представляет собой гибридный белок, состоящий из Fc-фрагмента Ig, связанного с CTLA4. Он применяется для ингибирования активации Т-клеток при ревматоидном артрите и трансплантации паренхиматозных органов.