Функциональная анатомия и физиология нервной системы - кратко с точки зрения внутренних болезней
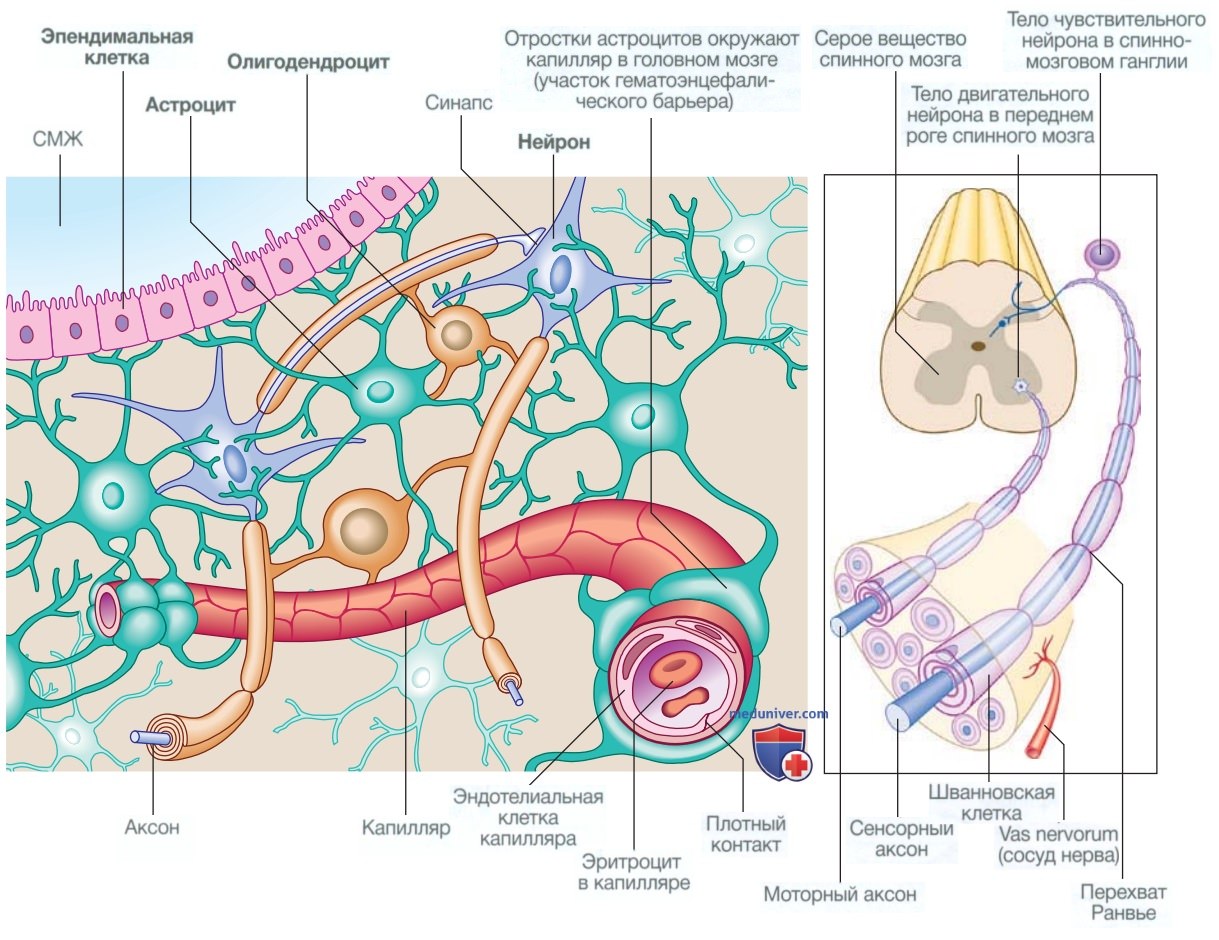
а) Клетки нервной системы. Нервная система состоит из миллиардов специализированных клеток, образующих впечатляющую сеть: головной мозг каждого человека имеет почти столько же связей, сколько песчинок существует во всем мире. Помимо нейронов, существуют три типа глиальных клеток.
Астроциты образуют структурную основу для нейронов и контролируют их биохимическую среду; отростки астроцитов примыкают к мелким кровеносным сосудам и формируют гематоэнцефалический барьер (рис. 1).
Рисунок 1. Клетки нервной системы. СМЖ — спинномозговая жидкость
Олигодендроциты отвечают за формирование и поддержание миелиновой оболочки, которая окружает аксоны и имеет большое значение для поддержания скорости и последовательности распространения потенциала действия вдоль аксонов. Периферические нервы представляют собой аксоны, заключенные в миелин, который вырабатывают олигодендроциты (шванновские клетки).
Микроглиальные клетки образуются из моноцитов/макрофагов и играют особую роль в борьбе с инфекцией и элиминации поврежденных клеток. Эпендимальные клетки выстилают желудочки головного мозга.
б) Генерация и передача нервного импульса. Роль центральной нервной системы (ЦНС) заключается в генерировании ответов на внешние раздражители и изменения внутреннего состояния. ЦНС должна поддерживать тонкий баланс между чувствительностью к внешним раздражителям и достаточной стойкостью, чтобы сохранять стабильность в быстро меняющейся среде.
Каждый нейрон получает входящие сигналы с помощью синаптической передачи от дендритов (разветвленных отростков других нейронов), которые суммируются, образуя выходящий сигнал в виде потенциала действия; затем потенциал действия проводится вдоль аксона и приводит к синаптической передаче на другие нейроны или на мышечные клетки в моторной системе.
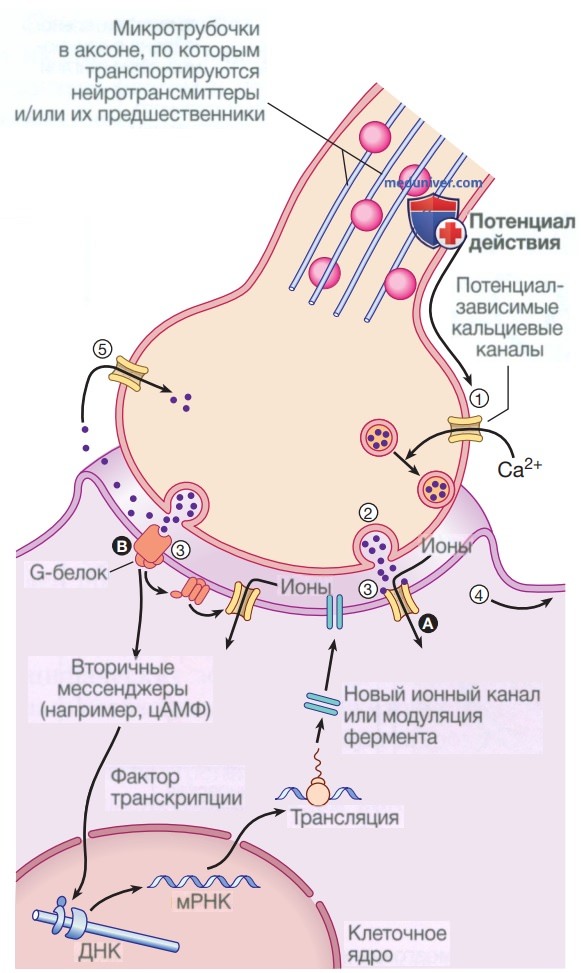
Суммирование входных сигналов обеспечивает совокупные изменения электрохимического градиента в нейроне-мишени, что при достаточной величине запускает потенциал действия. Связь между клетками осуществляется посредством синаптической передачи, которая задействует высвобождение нейротрансмиттеров для взаимодействия со структурами на поверхности клетки-мишени, включая ионные каналы и другие рецепторы на поверхности клетки (рис. 2).
Рисунок 2. Передача нервных импульсов и нейротрансмиттеры. (1) Потенциал действия, поступающий к нервному окончанию, деполяризует мембрану, и это открывает потенциал-зависимые кальциевые каналы. (2) Вход кальция в клетку вызывает слияние синаптических везикул, содержащих нейротрансмиттеры, с пресинаптической мембраной и высвобождение нейротрансмиттера в синаптическую щель. (3) Нейротрансмиттер связывается с рецепторами на постсинаптической мембране, (А) открывая лиганд-зависи-мые ионные каналы, которые запускают входящий ток ионов, деполяризуя мембрану и инициируя потенциал действия (4), или (В) связываясь с метаботропными рецепторами, которые активируют эффекторный фермент (например, аденилатциклазу) и, таким образом, модулируют транскрипцию гена через внутриклеточную систему вторичного мессенджера, что приводит к изменениям в синтезе ионных каналов или модуляции ферментов. (5) Нейротрансмиттеры попадают на пресинаптическую мембрану и/или метаболизируются. цАМФ — циклический аденозинмонофосфат; ДНК — дезоксирибонуклеиновая кислота; мРНК — рибонуклеиновая кислота
Известно, что по меньшей мере 20 различных нейротрансмиттеров действуют в разных местах нервной системы, и на большинство из них можно повлиять с помощью лекарственных препаратов.
Тело каждого нейрона может получать синаптические сигналы от тысяч других нейронов. Нервные окончания в синапсе также регулируются с помощью обратной связи через рецепторные участки на пресинаптической мембране, модифицируя высвобождение нейротрансмиттера через синаптическую щель. Помимо таких непосредственных эффектов, ряд нейротрансмиттеров обеспечивают длительную модуляцию метаболической функции или экспрессии генов.
Данный эффект, вероятно, лежит в основе более сложных процессов, таких как долговременная память.
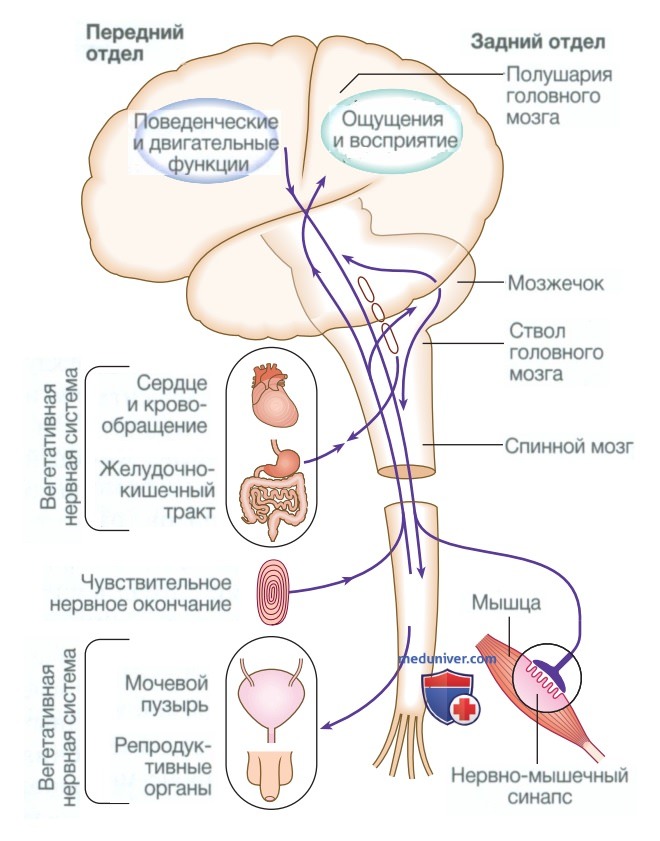
в) Функциональная анатомия нервной системы. Основные компоненты нервной системы и их взаимосвязи показаны на рис. 3.
Рисунок 3. Основные анатомические компоненты нервной системы
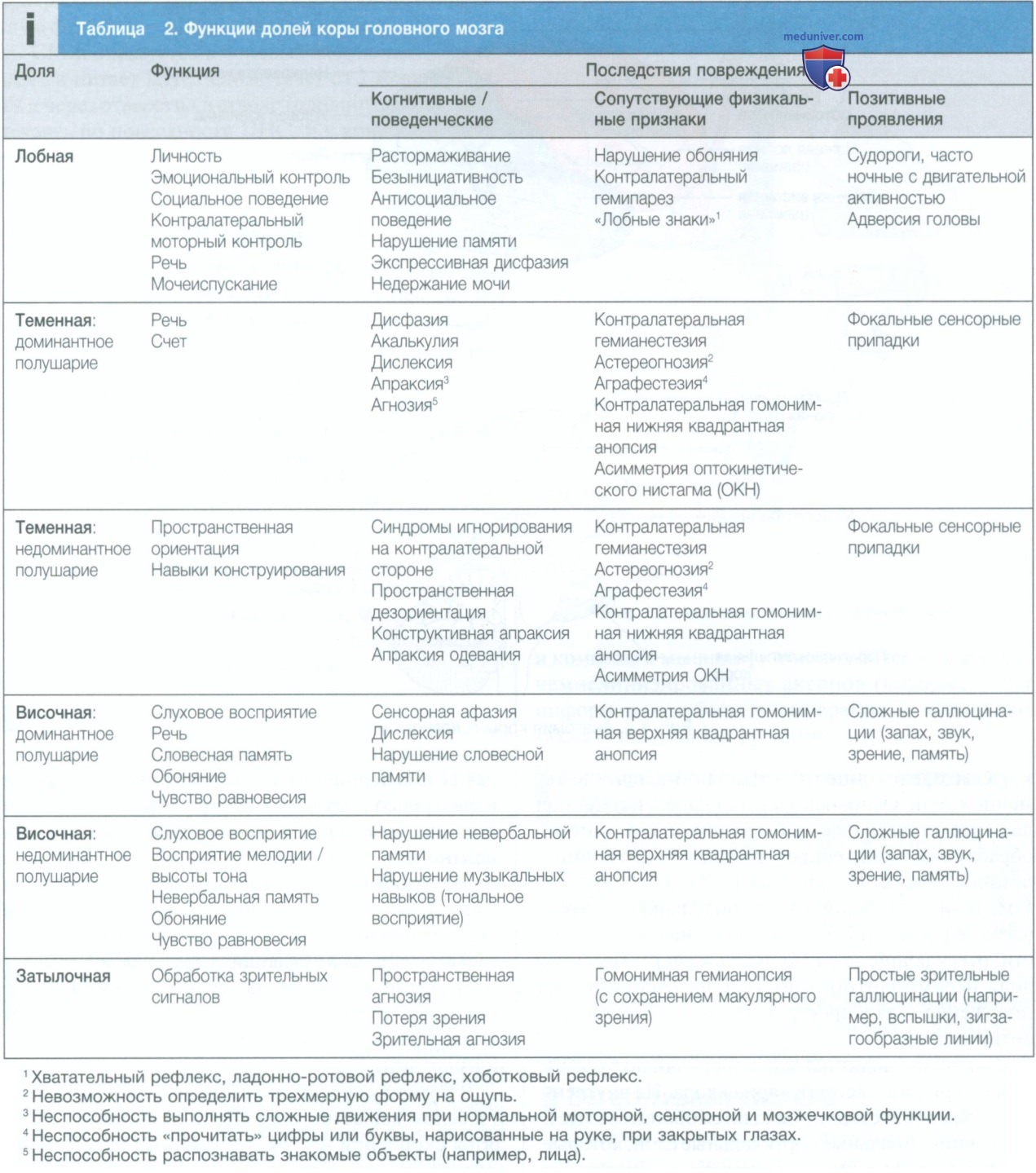
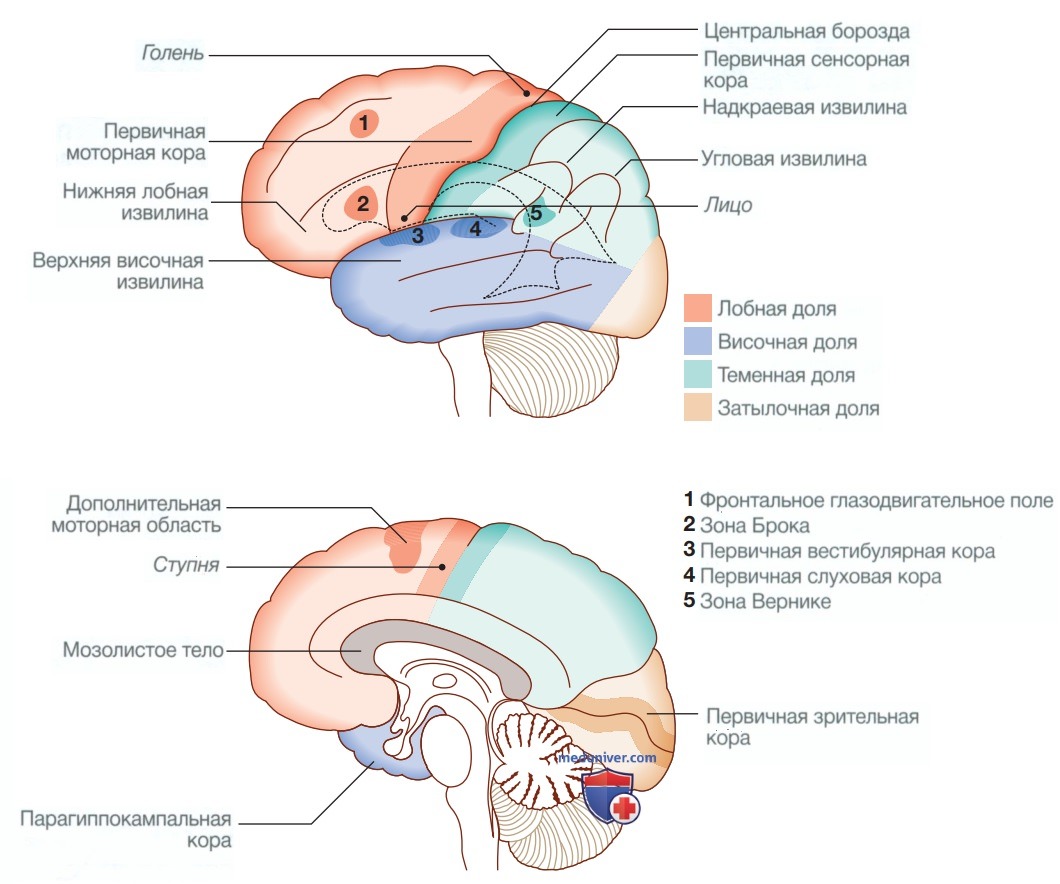
1. Полушария головного мозга. Полушария головного мозга координируют высший уровень нервной деятельности, передний отдел отвечает за исполнительные функции («делание»), а задний отдел формирует восприятие окружающей среды. Каждое полушарие головного мозга делится на четыре функционально специализированные доли (табл. 2 и рис. 4), причем некоторые функции распределены асимметрично («латерализованы»), чтобы обеспечить доминирование головного мозга для таких функций, как моторный контроль, речь и память.
Рисунок 4. Анатомия коры головного мозга
Доминирование одного из полушарий уравнивает доминирование конечностей с речевой функцией: у правшей левое полушарие почти всегда бывает доминантным, в то время как около половины левшей имеют доминантное правое полушарие.
Лобные доли отвечают за исполнительные функции, движение, поведение и планирование. Помимо первичной и дополнительной моторной коры, в них есть специализированные зоны, предназначенные для контроля движений глаз, речи (зона Брока) и мочеиспускания.
Теменные доли объединяют в одну целую сенсорную зону восприятия. Первичная сенсорная кора находится в постцентральной извилине теменной доли. Основная часть оставшейся теменной доли представлена «ассоциативной» корой, которая обрабатывает и интерпретирует данные различных органов чувств. Надкраевая и угловая извилины теменной доли доминантного полушария образуют часть речевой зоны. Рядом с ними находятся области, отвечающие за способность к счету. Теменная доля недоминантного полушария воспринимает положение тела в пространстве и отвечает за ориентирование.
В височных долях находится первичная слуховая кора и первичная вестибулярная кора. На внутренней медиальной стороне находятся обонятельные и парагиппокампальные кортикальные слои, которые участвуют в функции запоминания. Височные доли также тесно связаны с лимбической системой (включая гиппокамп и миндалину), участвующей в запоминании и эмоциональной обработке. Височная доля доминантного полушария также участвует в функциях речи, в частности в понимании устной речи (зона Вернике).
Обработка музыки происходит в обеих височных долях, ритма — на доминантной стороне, а мелодии и высоты тона — на недоминантной.
Затылочные доли отвечают за интерпретацию зрительных сигналов. Контралатеральная половина поля зрения представлена в каждой первичной зрительной коре, а окружающие области обрабатывают определенные зрительные субмодальности, такие как цвет, движение или глубина, и выполняют анализ более сложных зрительных структур (например, лиц).
В глубоких кортикальных слоях серого вещества и в белом веществе, состоящем из аксонов, находятся скопления клеток, называемые базальными ганглиями, которые отвечают за моторный контроль; таламус, отвечающий за уровень внимания к сенсорному восприятию; лимбическая система, связанная с эмоциями и памятью; и гипоталамус, отвечающий за гомеостаз (например, контроль температуры и аппетита). Желудочки головного мозга содержат СМЖ, которая выполняет амортизирующую функцию при движениях черепа.
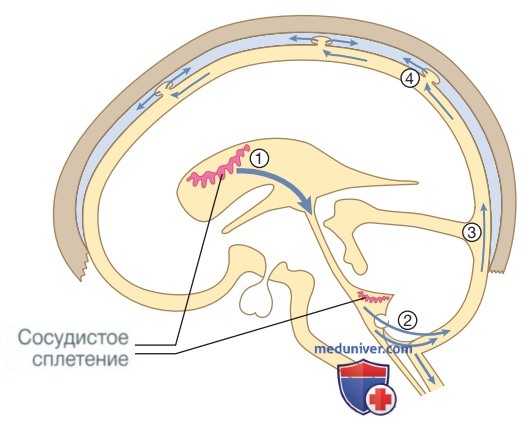
СМЖ образуется в боковых желудочках, защищает и питает ЦНС. СМЖ течет от I желудочка к IV и через отверстия в стволе головного мозга, растекаясь по поверхности ЦНС, и в конечном счете реабсорбируется в венозную систему головного мозга (см. рис. ниже).
Циркуляция спинномозговой жидкости. (1) Спинномозговая жидкость синтезируется в сосудистом сплетении желудочков и течет из бокового и III желудочков через водопровод в IV желудочек. (2) Через отверстия Люшки и Мажанди она выходит из головного мозга, растекается по полушариям (3) и стекает вниз по спинному мозгу и корешкам в субарахноидальном пространстве. (4) Затем она реабсорбируется в венозные синусы твердой мозговой оболочки через арахноидальные грануляции
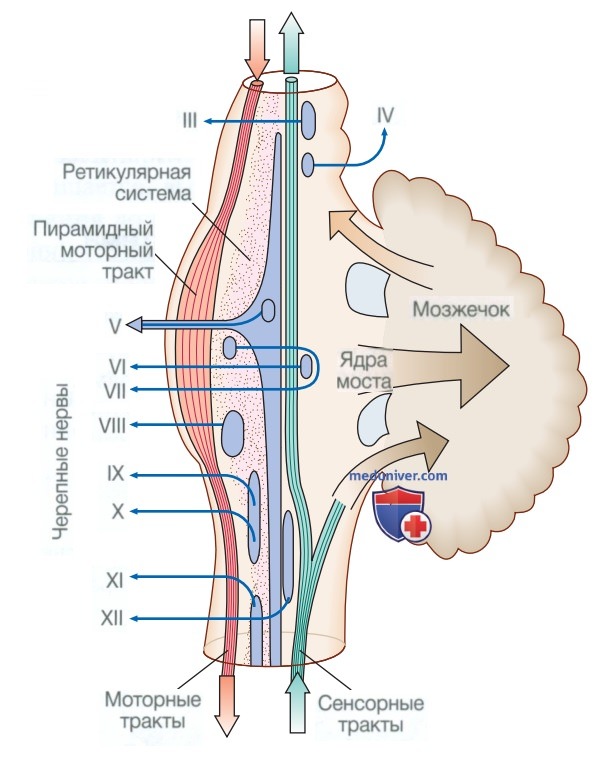
2. Ствол головного мозга. В стволе головного мозга проходят все сенсорные и моторные пути, входящие и выходящие из полушарий; кроме того, ствол головного мозга содержит ядра и проекционные волокна большинства черепных нервов, а также другие важные скопления нейронов в ретикулярной формации (рис. 5). Ядра черепных нервов обеспечивают двигательный контроль мышц головы (включая лицо и глаза) и координируют сенсорные сигналы от органов чувств и лица, носа, рта, гортани и глотки.
Рисунок 5. Анатомия ствола головного мозга
Они также передают вегетативные сигналы, включая функции зрачков, слюнных и слезных желез. Ретикулярная формация участвует преимущественно в контроле содружественных движений глаз, поддержании баланса и бодрствования, а также в регулировании сердечной деятельности и дыхания.
3. Спинной мозг. Спинной мозг — это путь практически для всех связей между внечерепными структурами и ЦНС. Афферентные и эфферентные волокна сгруппированы в отдельные пучки, а скопления клеток в сером веществе отвечают за моторные рефлексы низшего порядка и первичную обработку сенсорной информации.
4. Сенсорная периферическая нервная система. Тела чувствительных клеток периферических нервов расположены непосредственно за спинным мозгом — в спинномозговых ганглиях, находящихся в межпозвонковых отверстиях, в то время как дистальные части их нейронов используют различные специализированные окончания для преобразования внешних раздражителей в потенциалы действия.
Сенсорные (чувствительные) нервы состоят из комбинации толстых, быстрых, миелинизированных аксонов [которые несут информацию о проприоцепции (положении суставов) и команды к мышцам] и тонких, более медленных, немиелинизированных аксонов (которые несут информацию о боли и температуре и выполняют также вегетативные функции).
5. Моторная периферическая нервная система. Передние рога спинного мозга состоят из клеточных тел нижних мотонейронов. Для увеличения скорости проведения аксоны периферических двигательных нервов заключены в миелин, продуцируемый шванновскими клетками. Моторные (двигательные) нейроны высвобождают ацетилхолин в нервно-мышечный синапс, что приводит к изменению потенциала концевых пластинок и вызывает сокращение мышц.
6. Вегетативная система. Вегетативная система регулирует функцию сердечно-сосудистой и дыхательной системы, гладкие мышцы желудочно-кишечного тракта и множество экзокринных и эндокринных желез во всем организме. Вегетативная система централизованно управляется диффузными модулирующими системами в стволе головного мозга, лимбической системе, гипоталамусе и лобных долях, которые связаны с бодрствованием и основными поведенческими реакциями на угрозу.
Функционально и фармакологически сигналы вегетативной нервной системы делятся на два отдела: парасимпатическая и симпатическая система.
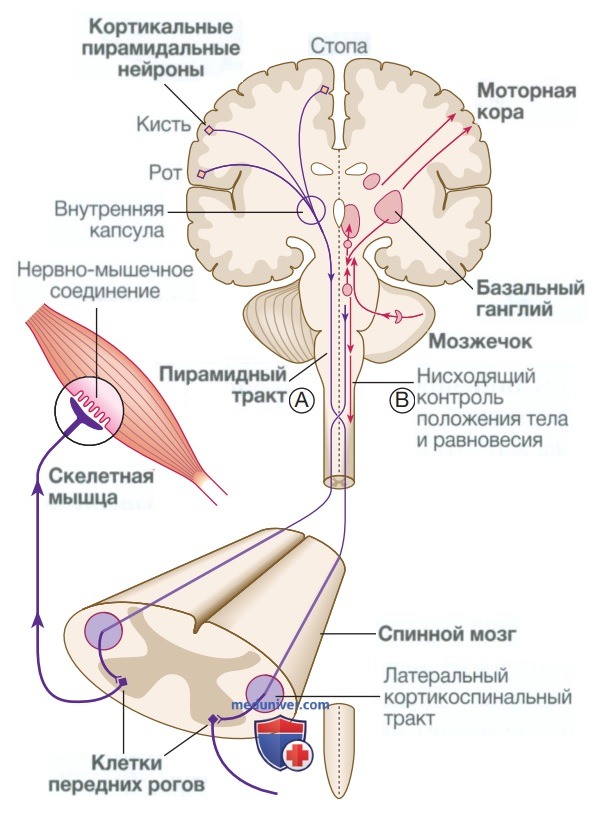
7. Моторная (двигательная) система. Программа движений, сформированная в премоторной коре, преобразуется в серию возбуждающих и тормозных сигналов в моторной коре, которые передаются в спинной мозг через пирамидный тракт (рис. 6). Он проходит через внутреннюю капсулу и вентральную часть ствола головного мозга, а затем переходит на другую половину (перекрест) в продолговатом мозге и входит в боковые столбы спинного мозга.
Рисунок 6. Моторная система. Отростки нейронов из моторной коры спускаются в виде пирамидного тракта во внутренней капсуле и ножке мозга к вентральной части ствола мозга, где большинство волокон переходит на противоположную сторону в нижней части продолговатого мозга (А). В спинном мозге верхние мотонейроны образуют кортикоспинальный тракт в боковом столбе спинного мозга перед образованием синапса с нижними мотонейронами в передних рогах спинного мозга. Активность в моторной коре регулируется базальными ганглиями и мозжечком. Пути, выходящие из этих структур, контролируют положение тела и равновесие (В)
«Верхние мотонейроны» пирамидного тракта образуют синапс с клетками передних рогов серого вещества спинного мозга, которые формируют нижние мотонейроны.
Любое движение требует изменения положения тела и мышечного тонуса, иногда в совершенно других мышечных группах, чем те, которые осуществляют фактическое движение. Моторная система состоит из иерархии органов управления, поддерживающих положение тела и мышечный тонус, на которые и накладывается любое движение. В сером веществе спинного мозга самый низкий порядок моторной иерархии отвечает за рефлекторные реакции на растяжение.
Мышечные веретена воспринимают удлинение мышцы; они обеспечивают афферентную сторону рефлекса растяжения и инициируют моносинаптический рефлекс, приводящий к защитному или реактивному сокращению мышц. Сигналы от ствола мозга оказывают в значительной степени тормозящее действие.
Полисинаптические связи в сером веществе спинного мозга контролируют более сложные рефлекторные сгибания и разгибания конечностей, которые образуют основные элементы скоординированных действий, но полный контроль требует сигналов от экстрапирамидной системы и мозжечка.
- Нижние мотонейроны. Нижние мотонейроны в передних рогах спинного мозга иннервируют группу мышечных волокон, называемых «моторной единицей». Утрата нижних мотонейронов приводит к невозможности мышечных сокращений внутри этой единицы, слабости и снижению мышечного тонуса. Впоследствии денервированные мышечные волокна атрофируются и самопроизвольно деполяризуются, вызывая фибрилляции.
За исключением языка, фибрилляции обычно заметны только на электромиографии (ЭМГ). С течением времени волокна соседних неповрежденных нейронов прорастают, обеспечивая повторную иннервацию, но нервно-мышечные синапсы увеличенных моторных единиц нестабильны и самопроизвольно деполяризуются, вызывая фасцикуляции (достаточно крупные, видимые невооруженным глазом). Следовательно, фасцикуляции указывают на хроническую денервацию с частичной повторной иннервацией.
- Верхние мотонейроны. Верхние мотонейроны оказывают как тормозящее, так и возбуждающее влияние на функцию нижних мотонейронов в передних рогах. Поражение верхних мотонейронов приводит к повышению мышечного тонуса, что хорошо заметно в наиболее сильных группах мышц (то есть в разгибателях нижних конечностей и сгибателях верхних конечностей).
Слабость при поражении верхних мотонейронов, наоборот, более выражена в противоположных мышечных группах. Утрата торможения также приводит к оживлению рефлексов и усилению рефлекторных движений, таких как отдергивание конечности в ответ на болевой раздражитель и спазм разгибателей. Повышенный тонус более выражен во время быстрого растяжения («спастическая фиксация»), но может быстро уступить место устойчивому напряжению (феномен «складного ножа»). Также появляются более примитивные рефлексы, такие как разгибательный подошвенный рефлекс.
Спастичность может развиться только через несколько недель после начала поражения верхних мотонейронов.
8. Экстрапирамидная система. Связи между базальными ганглиями и моторной корой составляют экстрапирамидную систему, которая контролирует мышечный тонус, положение тела и начало движения (см. рис. 6). Поражения экстрапирамидной системы вызывают увеличение тонуса, которое, в отличие от спастичности, сохраняется во всем диапазоне движений при любой скорости растяжения (ригидность по типу «свинцовой трубы»). Непроизвольные движения также являются признаком экстрапирамидных нарушений, а тремор в сочетании с ригидностью приводит к развитию характерной ригидности по типу «зубчатого колеса».
Экстрапирамидные нарушения также вызывают замедленные и неуклюжие движения (брадикинезия), размах которых обычно уменьшается при повторении, а также постуральную неустойчивость, способную спровоцировать падения.
9. Мозжечок. Мозжечок осуществляет тонкую настройку и координацию движения, инициируемого моторной корой, включая артикуляцию речи. Он также участвует в планировании и разучивании точных движений через реципрокные связи с таламусом и корой. Повреждение полушария мозжечка вызывает утрату координации на той же стороне тела. Дисфункция мозжечка нарушает плавность движений глаз, вызывая нистагм, и приводит к дизартричной речи. В конечностях начало движений происходит нормально, но при приближении к цели точность ухудшается, вызывая интенционный тремор.
Расстояния до целей оцениваются неверно (дисметрия), что приводит к «промахиванию». Также нарушается способность к быстрым, точным, регулярно чередующимся движениям (дисдиадохокинез). Червь мозжечка обеспечивает координацию походки и положения тела. Таким образом, нарушения в этой области вызывают характерную атактическую походку (см. ниже).
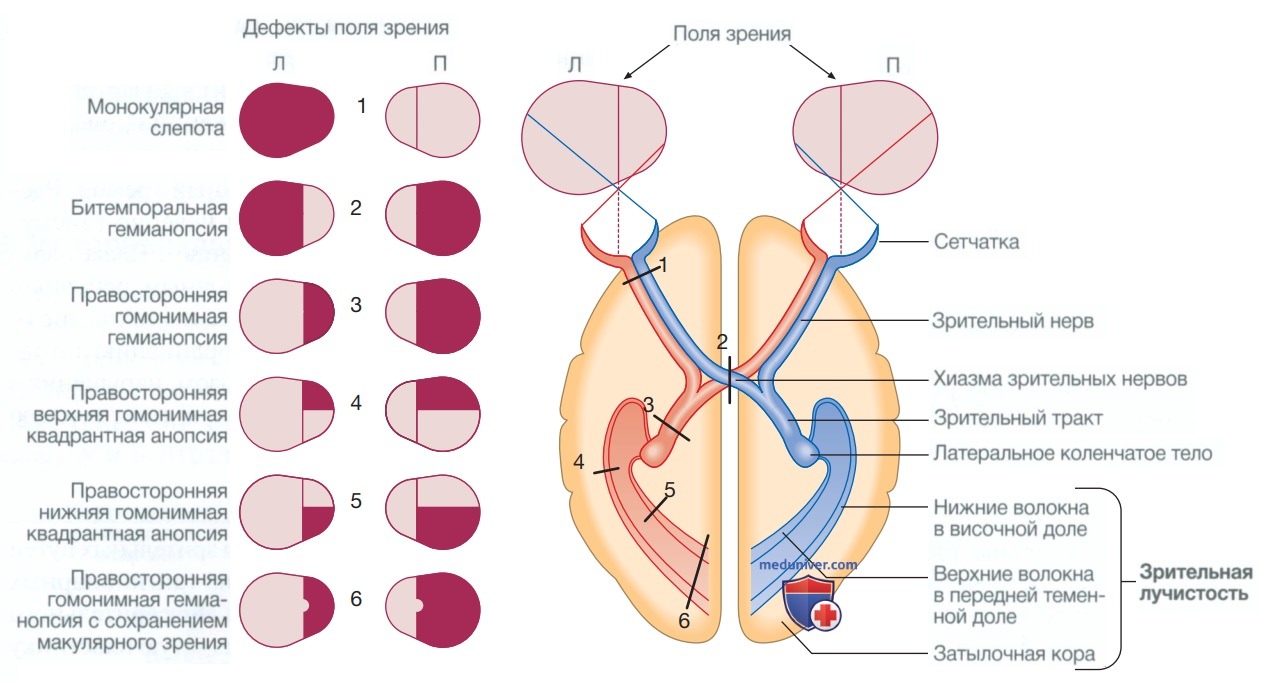
г) Зрение. Неврологическая организация зрительных путей показана на рис. 7. Волокна из ганглионарных клеток в сетчатке проходят через диск зрительного нерва, а затем обратно через решетчатую пластинку к зрительному нерву. Носовые волокна зрительного нерва (отвечающие за височное поле зрения) пересекаются в хиазме, а височные волокна — нет. Следовательно, волокна в каждом зрительном тракте и далее кзади несут картину контралатерального зрительного пространства. От латерального коленчатого ядра нижние волокна проходят через височные доли на пути к первичной зрительной области в затылочной коре, тогда как верхние волокна проходят через теменную долю.
Рисунок 7. Зрительные пути и дефекты полей зрения. Схематическое изображение глаз и головного мозга в поперечном разрезе
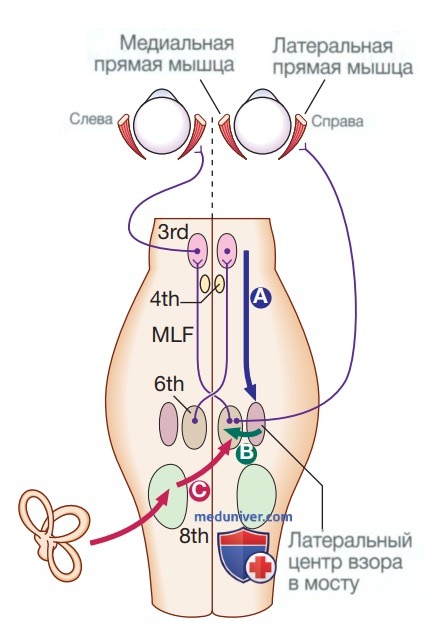
Обычно глаза движутся содружественно (в одном и том же направлении с одинаковой скоростью), хотя горизонтальная конвергенция позволяет объединять изображения на разных расстояниях. Контроль движений глаз начинается в полушариях головного мозга, в частности в области фронтальных глазодвигательных полей, а затем путь спускается к стволу мозга с входящими сигналами из зрительной коры головного мозга, верхнего холмика четверохолмия и мозжечка. Горизонтальные и вертикальные центры взора, расположенные в мосте и среднем мозге соответственно, координируют выходящие сигналы к ядрам глазодвигательного нерва (3, 4 и 6-му), которые связаны друг с другом медиальным продольным пучком (рис. 8).
Рисунок 8. Контроль содружественных движений глаз. Нисходящие проекционные волокна проходят от коры к латеральному центру взора в мосте (А). От центра взора отходят волокна к ядру VI черепного нерва (В), который иннервирует ипсилатеральную латеральную прямую мышцу и посылает проекционные волокна к контралатеральному ядру III нерва (и, следовательно, иннервирует медиальную прямую мышцу) через медиальный продольный пучок (МПП). Тонические сигналы от вестибулярного аппарата (С) попадают в контралатеральное ядро VI нерва через вестибулярные ядра
Медиальный продольный пучок особенно важен для координации горизонтальных движений глаз. Окончательные сигналы к внешним глазным мышцам поступают через глазодвигательный (III), блоковый (IV) и отводящий (VI) черепные нервы.
Размер зрачка определяется сочетанием парасимпатической и симпатической активности. Парасимпатические волокна берут начало в ядре Эдингера—Вестфаля III нерва и проходят вместе с III нервом до синапса в цилиарном ганглии перед иннервацией сфинктера зрачка в радужной оболочке. Симпатические волокна начинаются в гипоталамусе, проходят через ствол головного мозга и шейный отдел спинного мозга, появляются в Т1, возвращаются к глазу вместе с внутренней сонной артерией и иннервируют дилататор зрачка.
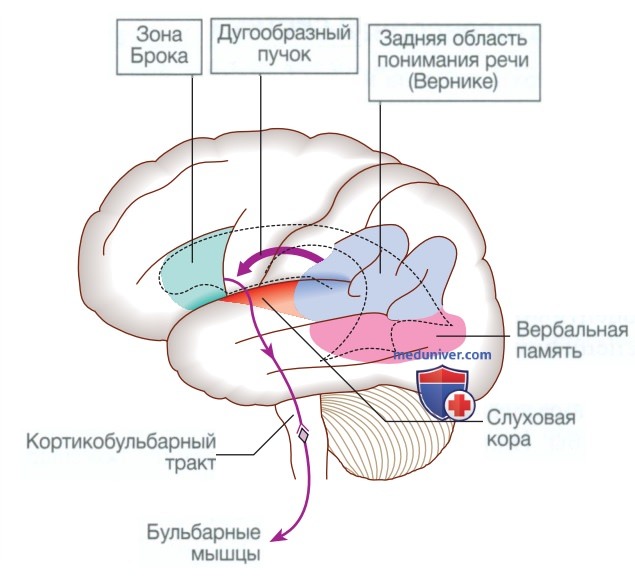
д) Речь. Значительная часть коры головного мозга участвует в процессе формирования и интерпретации речевых сообщений, особенно в доминантном полушарии (см. табл. 2). Расшифровка речевых звуков (фонем) осуществляется в верхней части задней височной доли. Определение значения, а также формулировка речи, необходимая для выражения идей и концепций, происходит преимущественно в нижних частях передней теменной доли (угловая и надкраевая извилины). Область восприятия речи в височной доле называется зоной Вернике (рис. 9). Другие части височной доли участвуют в формировании вербальной памяти, где «хранятся» лексиконы значимых слов.
Рисунок 9. Области коры головного мозга, участвующие в генерации разговорной речи
Части теменной доли недоминантного полушария, по-видимому, участвуют в невербальных аспектах языка, распознавании значимых интонационных паттернов (просодия).
Лобная зона речи находится в заднем конце нижней лобной извилины доминантного полушария, известной как зона Брока. Она получает сигналы от височных и теменных долей через дугообразный пучок. Моторные команды, генерируемые в зоне Брока, поступают к ядрам черепных нервов в мосту и в продолговатом мозге, а также к клеткам передних рогов спинного мозга. Нервные импульсы к губам, языку, нёбу, глотке, гортани и дыхательным мышцам приводят к формированию серии упорядоченных звуков, составляющих речь. Мозжечок также играет важную роль в координации речи, а поражение мозжечка приводит к дизартрии, при которой нарушается моторная артикуляция речи.
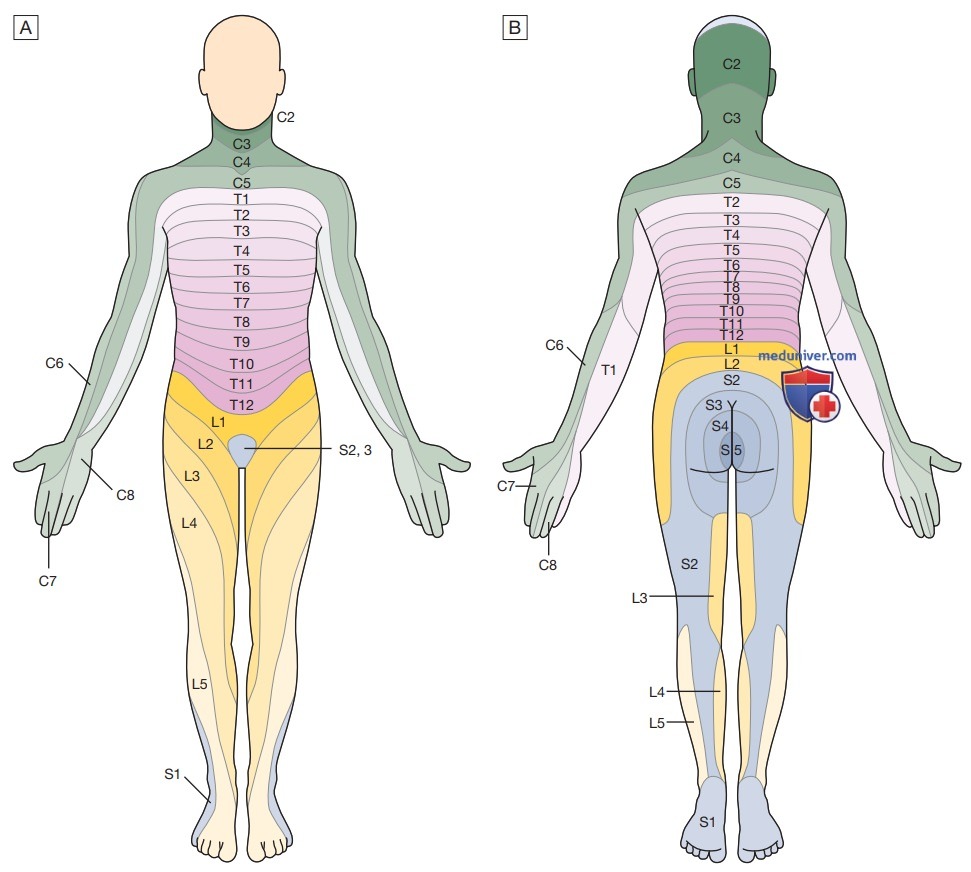
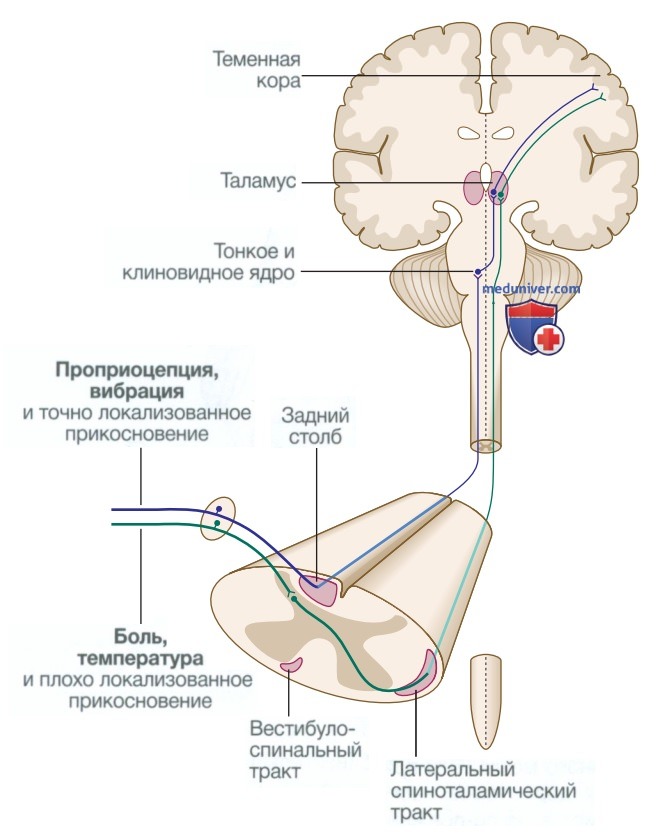
е) Соматосенсорная система. Поверхность тела может быть описана с помощью дерматомов, где каждый дерматом представляет собой участок кожи, в котором чувствительные (сенсорные) нервы выходят от одного спинномозгового корешка (рис. 10). Сенсорная информация передается в двух анатомически отдельных восходящих системах (рис. 11). Волокна от проприоцептивных органов и волокна, которые передают специфические ощущения (включая вибрацию), входят в спинной мозг через задние рога и проходят без образования синапса в ипсилатеральные задние столбы.
Рисунок 10. Участки, иннервируемые определенными отделами спинного мозга. Это приблизительные границы, и в реальности они значительно перекрываются. Клиническая польза этих дерматомов несколько уменьшилась с появлением высококачественной магнитно-резонансной томографии спинного мозга, но по-прежнему важно установить наличие «спинального уровня» ощущений, запомнить иннервацию седловидной области и отметить нисходящую траекторию некоторых лицевых спиноталамических путей в шейном отделе. Вид спереди (А). Вид сзади (В)
Рисунок 11. Основные соматические сенсорные пути
Напротив, волокна, передающие болевые и температурные ощущения (ноцицептивные нейроны), образуют синапсы с нейронами второго порядка, которые пересекают срединную линию в спинном мозге перед переходом в контралатеральный переднелатеральный спиноталамический тракт к стволу головного мозга.
Нейроны второго порядка заднего столба сенсорной системы пересекают срединную линию в верхней части продолговатого мозга и поднимаются через ствол головного мозга. Здесь они расположены медиально относительно (уже пересеченного) спиноталамического пути. Поэтому поражение ствола головного мозга может привести к потере всех видов чувствительности на противоположной стороне тела. Распределение потери чувствительности на лице из-за повреждения ствола головного мозга зависит от анатомии волокон тройничного нерва в стволе головного мозга.
Волокна от задней части лица (около ушей) спускаются в стволе головного мозга до верхней части спинного мозга перед образованием синапса; аксоны нейронов второго порядка пересекают срединную линию и затем поднимаются вместе со спиноталамическими волокнами. Волокна, передающие ощущения от передних областей лица, спускаются на меньшее расстояние в стволе головного мозга. Таким образом, потеря чувствительности на лице из-за повреждений нижней части ствола головного мозга распределяется в виде «балаклавы», так как поражаются наиболее длинные нисходящие волокна тройничного нерва. Спинной и спиноталамический тракты заканчиваются в таламусе, будучи перенаправленными оттуда в теменную кору.
- Боль. Боль — это сложное ощущение, которое только частично связано с активностью ноцицептивных нейронов. В высших отделах хроническая и сильная боль интенсивно взаимодействует с настроением и может усугубиться расстройством настроения, включая депрессию и тревогу. Модификация психологических и психиатрических последствий является очень важной частью лечения боли.
- Контроль сфинктеров. Симпатическая иннервация мочевого пузыря исходит из корешков Т11—L2 до синапса в нижнем подчревном сплетении, в то время как парасимпатическая иннервация исходит из S2-4. Кроме того, внешний (произвольный) сфинктер получает соматическую иннервацию из S2_4, через половые (срамные) нервы.
Накопление мочи обеспечивается путем подавления парасимпатической активности и, таким образом, расслабления мышцы детрузора в стенке мочевого пузыря. Удержанию мочи также помогает одновременное тоническое сокращение уретральных сфинктеров, опосредованное симпатической и соматической иннервацией. Опорожнение мочевого пузыря у взрослых обычно происходит осознанно, с ослаблением тонического торможения в центре мочеиспускания в мосте под влиянием более высоких центров, что приводит к расслаблению мышц тазового дна и наружного и внутреннего уретральных сфинктеров, наряду с опосредованным парасимпатической иннервацией сокращением детрузора.
ж) Личность и настроение. Физиология и патология расстройств настроения (синоним — аффективные расстройства) обсуждаются в другой главе, но важно помнить, что любой процесс, влияющий на функцию мозга, может влиять на настроение и эмоциональное состояние. И наоборот, расстройство настроения может оказать существенное воздействие на восприятие и функцию. Нередко бывает сложно определить, являются ли психологические и психические изменения причиной или следствием каких-либо неврологических симптомов.
- Сон. Функция сна неизвестна, но она необходима для здоровья. Сон контролируется активирующей ретикулярной системой в верхнем отделе ствола головного мозга и промежуточном мозге. Он состоит из разных этапов, которые можно визуализировать с помощью электроэнцефалограммы (ЭЭГ). Когда появляется сонливость, нормальный фоновый альфа-ритм ЭЭГ исчезает, и в активности преобладают глубокие медленные волны. По мере того как сон углубляется и начинаются сновидения, конечности становятся вялыми, движения «блокируются» и на медленные волны накладываются ЭЭГ-признаки быстрых движений глаз (REM).
REM-сон длится короткое время, прежде чем начинается другой период — медленноволнового сна; этот цикл повторяется несколько раз в течение ночи. Фазы REM удлиняются по мере развития сна. REM-coh, по-видимому, является наиболее важной частью цикла сна для освежения когнитивных процессов, а лишение REM-сна вызывает усталость, раздражительность и нарушение формирования суждений.
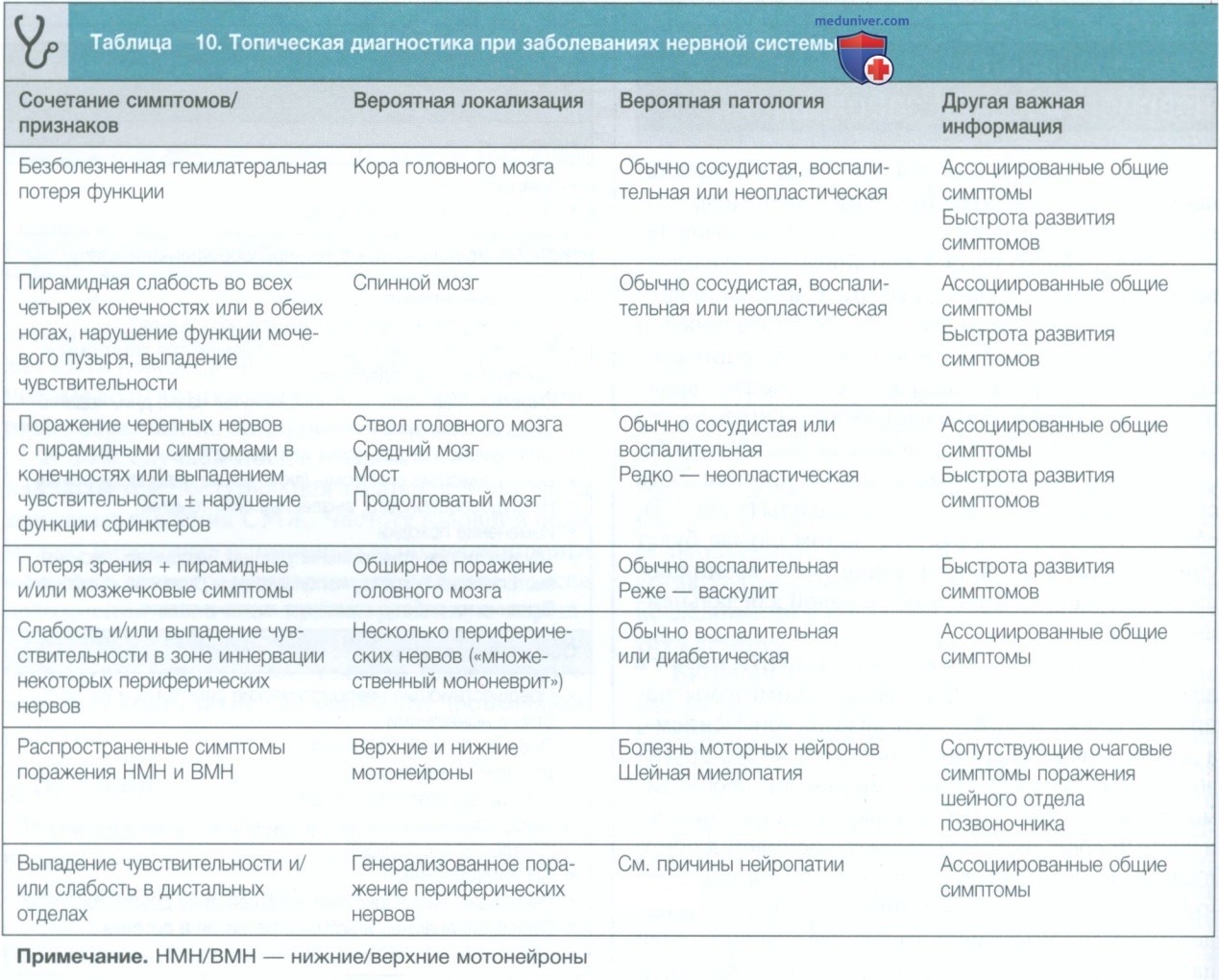
з) Определение места поражения в центральной нервной системе. После сбора анамнеза и осмотра пациента клиницист должен сформировать представление о характере и локализации патологии (см. табл. 10). Учитывая сложную анатомию ствола головного мозга, в этом разделе мы остановимся на возможной локализации более подробно (см. рис. 5).
Поражения ствола головного мозга, как правило, характеризуются симптомами, связанными с дисфункцией черепного нерва, мозжечка и верхних мотонейронов, и чаще всего вызваны сосудистыми заболеваниями. Поскольку анатомия ствола головного мозга очень точно организована, обычно можно локализовать место поражения на основании тщательного сбора анамнеза и осмотра, чтобы точно определить, какие участки/ядра поражены, обычно при минимальном количестве очагов.
Например, у пациента с внезапным появлением симптомов нарушения функции верхних мотонейронов, затрагивающих правую часть лица и правые руку и ногу в сочетании с левосторонним параличом III нерва, очаг поражения будет в левой ножке мозга в стволе головного мозга, и, вероятно, причиной окажется очаговый инсульт, так как появление симптомов было внезапным. Эта комбинация симптомов и признаков известна как синдром Вебера и является одним из нескольких подробно описанных стволовых альтернирующих синдромов, которые перечислены в табл. 3.
Влияние дефицитов отдельных черепных нервов обсуждается в разделах, посвященных движениям глаз и слабости лицевых мышц, потере чувствительности при повреждениях ствола головного мозга, дисфонии и дизартрии и бульбарным симптомам.