Функциональная анатомия и физиология боли - кратко с точки зрения внутренних болезней
Боль — это неприятное сенсорное и эмоциональное ощущение, связанное с фактическим или потенциальным повреждением ткани или описываемое в терминах такого повреждения. Это одна из самых распространенных жалоб, с которой люди обращаются за медицинской помощью.
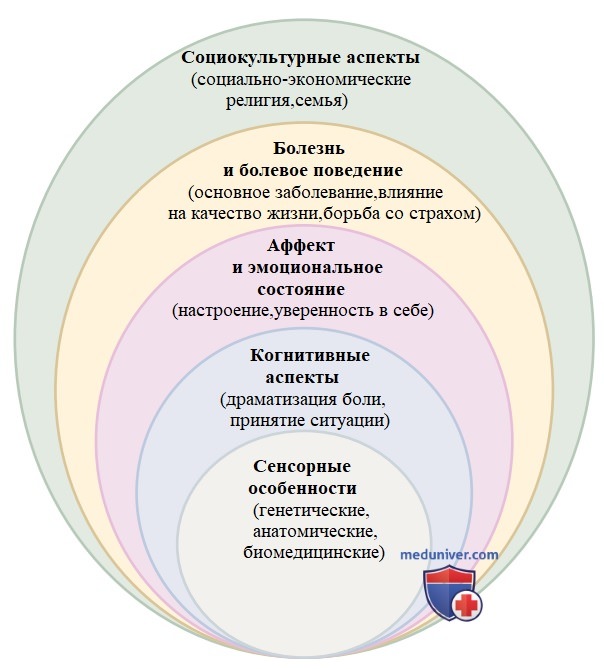
Наше понимание механизмов боли в значительной степени развилось из предположения Гиппократа в 450 г. до н.э., что боль возникает в результате дисбаланса жизненно важных жидких сфер организма. В настоящее время известно, что боль — это сложный симптом, на который влияют (и модифицируют его) многие социальные, культурные и эмоциональные факторы (рис. 1).
Рисунок 1. Биопсихосоциальная модель боли. Восприятие боли как симптома зависит не только от сенсорных воздействий, но и от когнитивной реакции человека на боль, его эмоционального состояния, основного заболевания, а также социального и культурного уровня
Ощущение острой боли, возникающей в ответ на воспаление или повреждение ткани, играет важную роль в защите от дальнейшего повреждения. Хроническая боль не выполняет никакой полезной функции, но приводит к значительным страданиям пациента и имеет большое социальное значение.
Далее обсуждаются механизмы и медиаторы, которые участвуют в формировании боли.
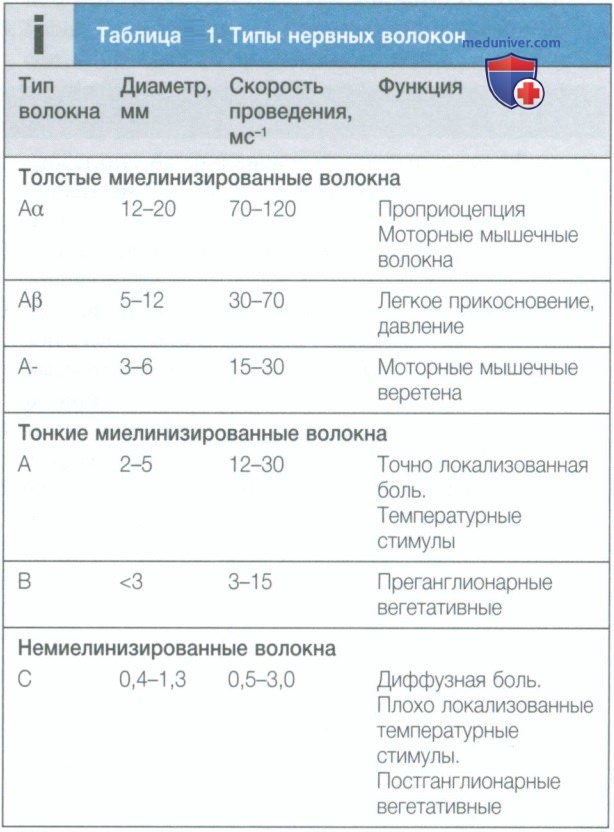
а) Периферические нервы. Периферические нервы образованы несколькими типами нейронов. Их можно разделить на две группы в зависимости от наличия у них миелиновой оболочки. Миелинизированные волокна имеют быструю скорость проведения и отвечают за передачу различных сенсорных сигналов, таких как проприоцепция, легкое прикосновение, тепло и холод, а также за обнаружение локализованных болевых стимулов, например от укола.
Немиелинизированные волокна имеют гораздо более медленную скорость проведения и отвечают за передачу диффузной и слабо локализованной боли, а также других ощущений (табл. 1).
Сенсорные нейроны (также называемые первичными афферентными нейронами) соединяют спинной мозг с периферией и иннервируют определенную территорию или дерматом, что можно использовать для определения места поражения нерва. У здоровых людей дерматомы имеют четкие границы, но при патологических болевых синдромах они могут стать размытыми в результате нейропластичности — ощущение боли в области, смежной с той, которая иннервируется определенным нервным окончанием.
Вегетативные нейроны также содержат болевые волокна и отвечают за передачу висцеральных ощущений, таких как колики. В целом висцеральная боль диффузная и локализована хуже, чем боль, передаваемая сенсорными нейронами.
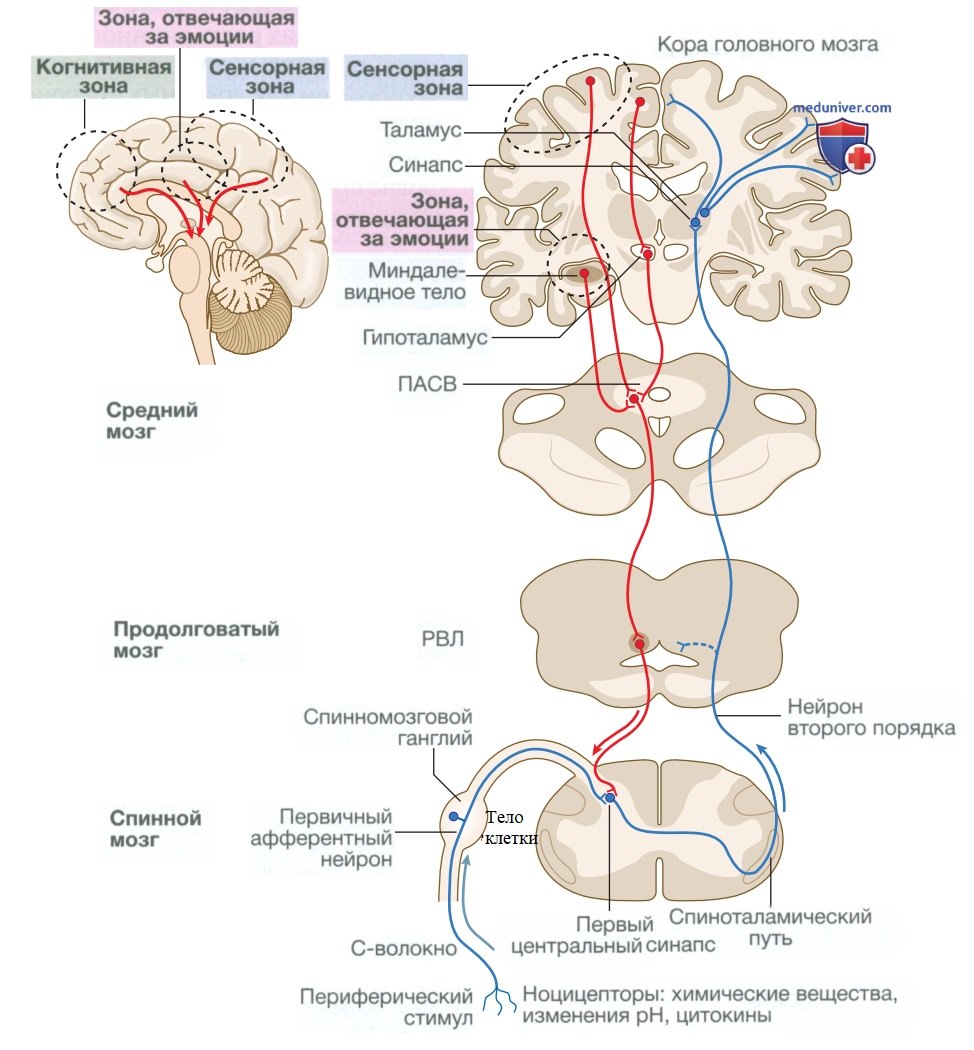
Анатомические особенности афферентного болевого пути показаны на рис. 2. Болевые сигналы передаются от периферии к спинному мозгу с помощью сенсорных нейронов, которые состоят:
Рисунок 2. Восходящие и нисходящие болевые пути. Восходящие пути изображены синим цветом, нисходящие — красным. Болевые сигналы воспринимаются на периферии ноцицепторами, которые активируются химическими веществами, изменениями pH и цитокинами. Сигнал передается первичным афферентным нейроном в спинной мозг, где находится синапс с нейроном второго порядка, который далее передает сигнал к таламусу. После этого болевой сигнал передается в кору головного мозга. Интенсивность болевых сигналов подвергается обширной модуляции на нескольких уровнях в нервной системе. Когнитивные воздействия от лобной доли в сочетании с сенсорными влияниями коры и эмоциональными эффектами миндалевидного тела влияют на восприятие боли в среднем мозге вокруг периакведуктального серого вещества и ростровентролатерального отдела в продолговатом мозге. Эти структуры образуют часть нисходящих модулирующих систем, которые в норме препятствуют восприятию боли. Однако при некоторых хронических болевых синдромах может возникнуть дисфункция нисходящих путей, усиливающая боль
• из тела клетки, содержащего ядро и расположенного в спинномозговом ганглии вблизи спинного мозга. Тело клетки необходимо для выживания нейрона, образования нейротрансмиттеров и функции нейронов;
• нервного волокна (аксона) и периферических нервных окончаний, располагающихся на периферии и содержащих ряд рецепторов в нейрональной мембране;
• специализированных рецепторов на периферии, состоящих из голых нервных окончаний, известных как ноцицепторы, или болевые рецепторы, которые активируются различными медиаторами. Они располагаются преимущественно в эпидермисе;
• центрального окончания, которое подходит к заднему рогу спинного мозга, образуя первый центральный синапс с нейронами, передающими болевые ощущения в головной мозг.
При встрече с болевым стимулом активация ноцицепторов приводит к генерации потенциала действия, который распространяется вверх к спинномозговому ганглию, а также стимулирует высвобождение нейротрансмиттеров, оказывающих вторичное воздействие на окружающие нейроны.
б) Спинной мозг. Сенсорные нейроны посредством центральных окончаний образуют синапс с нейронами второго порядка в задних рогах спинного мозга. Здесь происходит значительная модуляция болевых стимулов под действием как локальных нейронов в спинном мозге, так и нисходящих путей нейронов из головного мозга.
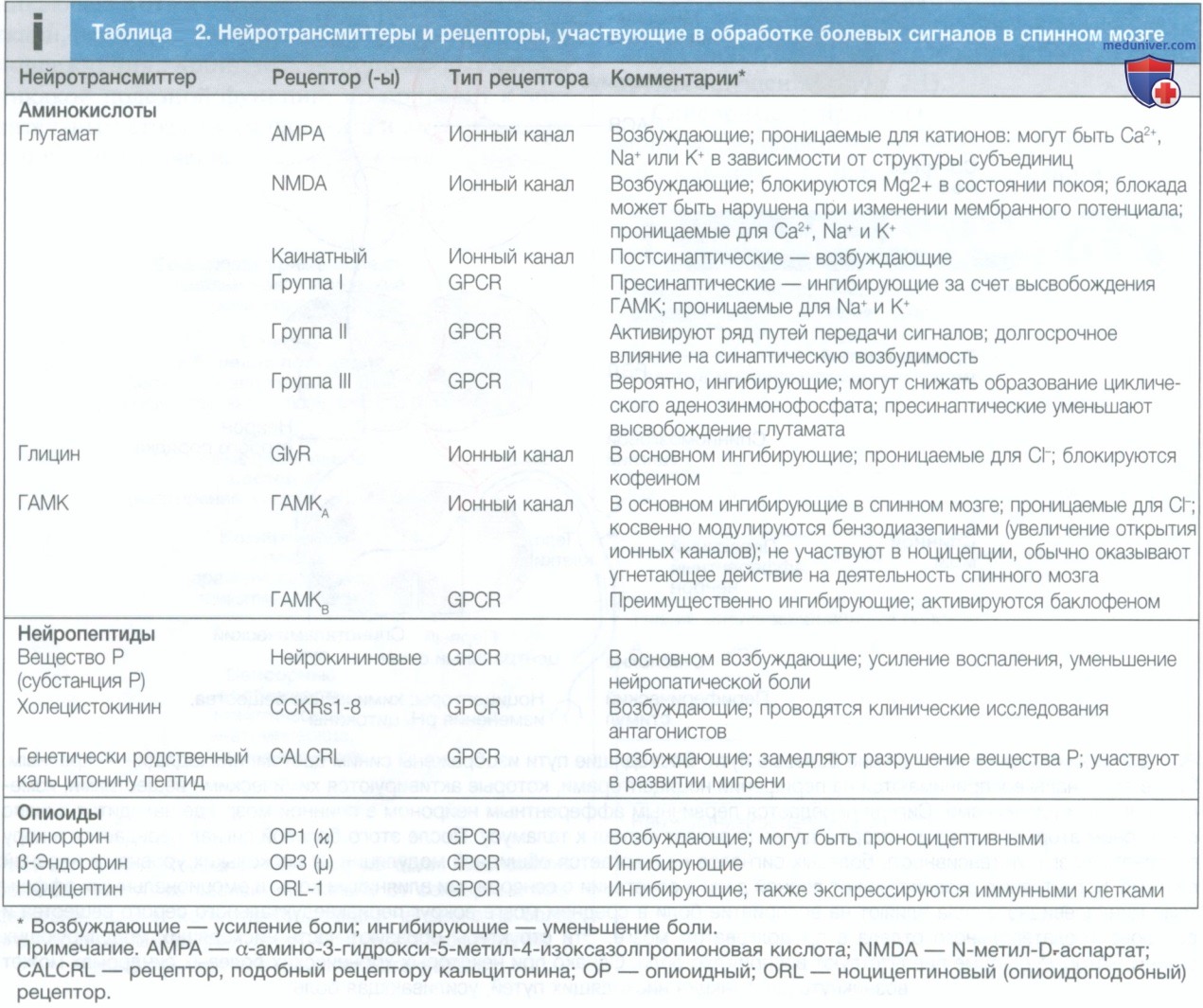
В обработке болевых сигналов на этом уровне принимает участие ряд нейротрансмиттеров, представленных в табл. 2. К ним относят аминокислоты, такие как глицин и ГАМК, являющиеся тормозными нейромедиаторами, и глутамат, являющийся возбуждающим медиатором; нейропептиды, такие как вещество Р и генетически родственный кальцитонину пептид (calcitonin gene-related peptide), и эндорфины. Увеличивают ли они или уменьшают восприятие боли — зависит от связи с нейронами, на которые они воздействуют.
в) Центральная обработка (процессинг) боли. Сигналы, передаваемые нейронами второго порядка в спинном мозге, доставляются в сенсорную кору нейронами третьего порядка, которые образуют синапсы с нейронами второго порядка в таламусе. Здесь на восприятие боли влияют взаимодействия ряда структур в головном мозге, в которых задействованы сенсорные, когнитивные и эмоциональные аспекты. Все вышеперечисленное носит название «нейроматрицы боли» (см. рис. 2).
Сигналы внутри нейроматрицы имеют разнонаправленную природу, включая модуляцию входящих сообщений корой головного мозга (нисходящая регуляция), а также сложную сеть взаимосвязей между другими подкорковыми структурами. В норме наблюдается определенная степень нисходящего торможения со стороны ствола головного мозга, которая уменьшает импульсы от периферических раздражителей.
Считается, что хроническая генерализованная боль и опиоидиндуцированная гипералгезия могут быть вызваны, по крайней мере отчасти, нарушением центральной обработки болевых сигналов. Высказывалось также предположение о том, что из-за различий в уровнях нисходящего торможения у некоторых лиц предрасположенность к развитию хронической боли может быть выше. В последние годы возрастает интерес к роли глиальных клеток в обработке болевых сигналов.
Как астроциты, так и клетки микроглии могут активироваться при хронических болевых состояниях и выделять провоспалительные цитокины, а также изменять обратный захват возбуждающих нейротрансмиттеров, таких как глутамат, которые могут значительно влиять на восприятие боли. По мере улучшения понимания этих процессов возрастает потенциал для разработки новых методов лечения, действующих на данные медиаторы, и по данному направлению уже проведены начальные фазы ряда клинических исследований у пациентов с нейропатической болью.
г) Сенситизация. Сенситизация является одним из основных явлений при обработке болевых сигналов. Считается, что как периферическая, так и центральная нервная система быстро адаптируются к боли, особенно в ответ на повреждение тканей. Этот адаптационный процесс называется нейропластичностью. В некоторых ситуациях нейропластичность может привести к долгосрочным изменениям путей, участвующих в восприятии и обработке ноцицептивных стимулов, что приводит к хроническим болевым синдромам.
Специфические изменения в основных нейротрансмиттерах и рецепторах различаются при хронических болевых состояниях, что влияет на эффективность лечения. Например, экспрессия μ-опиоидных рецепторов подавляется при нейропатической боли, что может привести к снижению эффективности опиоидных анальгетиков.
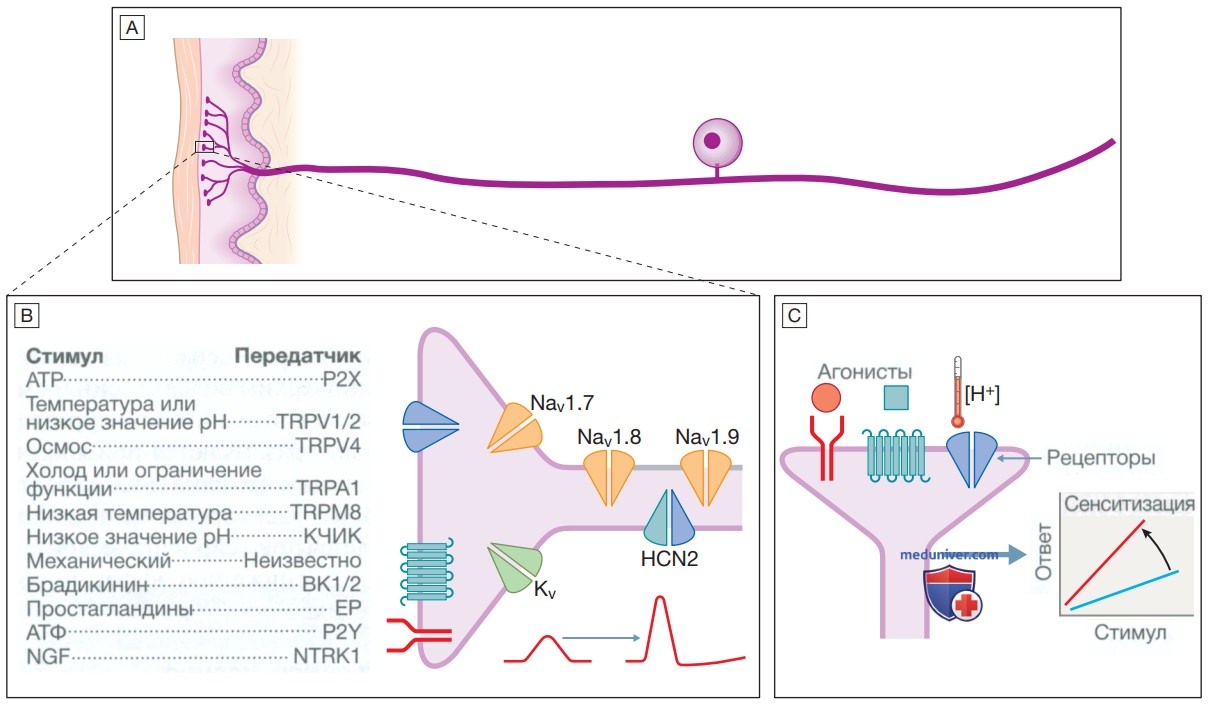
1. Периферическая сенситизация. Периферическая сенситизация может возникать при различных клинических состояниях, включая сепсис, рак, воспалительные заболевания, травмы, хирургическое вмешательство и ожирение. Финальным общим механизмом сенситизации для всех этих состояний является воспаление. Воспаление сопровождается повышенной проницаемостью капилляров и отеком тканей с выделением различных медиаторов, включая брадикинин, ионы водорода, простагландины и аденозин, которые связываются с рецепторами и ионными каналами на ноцицепторах первичных афферентных нейронов (рис. 3).
Рисунок 3. Механизмы периферической сенситизации. А — сенсорный нерв, оканчивающийся ноцицептором в коже. В — периферические ноцицепторы экспрессируют различные рецепторы и ионные каналы, которые действуют как медиаторы боли. Они включают натриевые каналы, участвующие в патогенезе врожденных болевых синдромов; пуринергический 2Х (Р2Х) рецептор аденозинтрифосфата; члены надсемейства ионных каналов с транзиторным рецепторным потенциалом (transient receptor potential — TRP), которые определяют изменения осмоляльности и температуры; кислоточувствительные ионные каналы, которые определяют ионы водорода; рецепторы, сопряженные с G-белком, которые определяют брадикинин, простагландины и аденозинтрифосфат; и рецептор нейротрофической тирозинкиназы 1 (neurotrophic tyrosine kinase 1), который обнаруживает фактор роста нервов (nerve growth factor). С — активация этих рецепторов лигандами, ионами водорода [Н+] и высокой температурой (>42 °C) увеличивает потенциалы действия, которые усиливают болевые сигналы и вызывают периферическую сенситизацию. С изменениями по: Bennett D.L., Woods C.G. Painful and painless channelopathies.
Пути передачи сигналов, активируемые этими медиаторами, генерируют потенциалы действия, которые передаются сенсорными нейронами в спинной мозг. Если вызывающие боль стимулы сохраняются, порог активации сенсорных нейронов уменьшается, что приводит к усилению передачи болевых сигналов в спинной мозг.
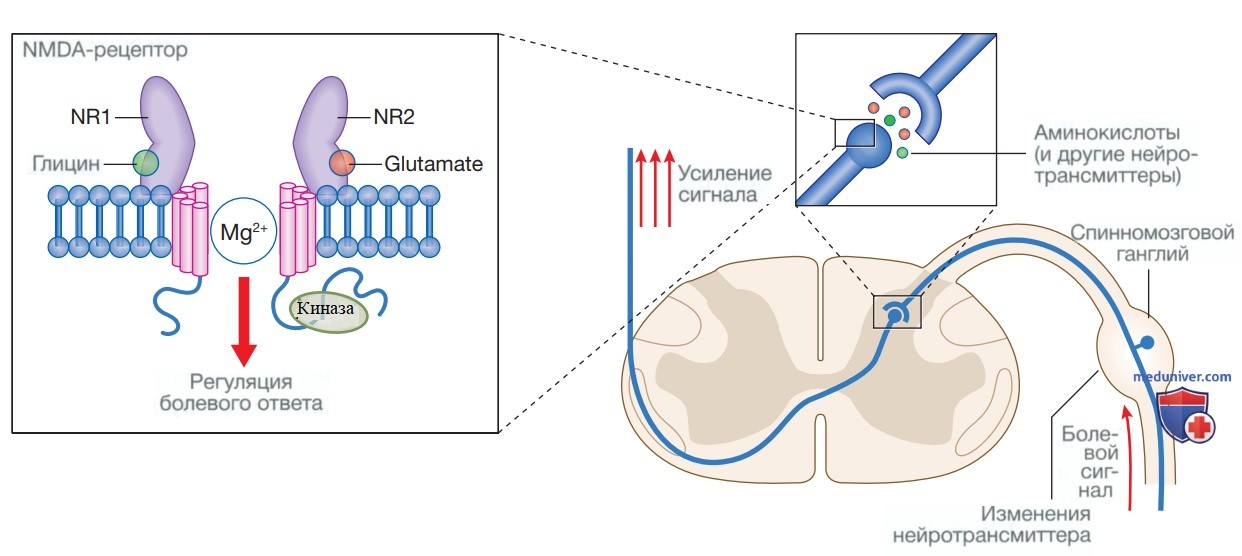
2. Центральная сенситизация. Сенситизация также может происходить на уровне спинного мозга в ответ на устойчивый болевой стимул. Она может быть острой и быстрой (например, в ранний послеоперационный период) или прогрессировать и хронизироваться в условиях хронической инфекции, при раке, повторных операциях или множественных травмах. Глутамат, действуя через рецепторный комплекс N-метил-D-аспартат (N-methyl-D-aspartate), играет основную роль в центральной сенсибилизации (рис. 4).
Рисунок 4. Механизмы центральной сенситизации. Для постсинаптической активации рецептора N-метил-O-аспартата требуется наличие аминокислот глицина и глутамата, которые связываются с субъединицами NR1 и NR2 соответственно; они усиливают болевые сигналы на уровне спинного мозга. Напротив, ионы магния блокируют активацию данного рецептора
В ответ на продолжительный периферический болевой стимул в спинном мозге высвобождается повышенное количество глутамата, перекрывающее ингибирующее действие ионов магния, что приводит к активации рецептора N-метил-D-аспартата. Это инициирует каскад внутриклеточных путей передачи сигналов, вследствие чего происходит долгосрочная модификация соматосенсорной обработки стимулов с усилением болевых реакций в спинном мозге и сохранением возбуждения нейронов даже после исчезновения болевого стимула. Это явление называется разрядом последействия.
При нейропатической боли длительная активация пути N-метил-D-аспартата приводит к уменьшению количества ингибирующих промежуточных нейронов, что еще больше усиливает боль.
д) Генетические детерминанты восприятия боли. В реакции людей на болевые стимулы отмечаются существенные этнические и индивидуальные различия. Исследования у близнецов показали, что наследуемость хронической генерализованной боли составляет от 30 до 50%. В общей популяции индивидуальные варианты ответа на боль и восприятия боли наиболее вероятно обусловлены сложным взаимодействием между генетическими и внешнесредовыми факторами. В ряде случаев были получены достоверные взаимосвязи между генотипом и хронической генерализованной болью.
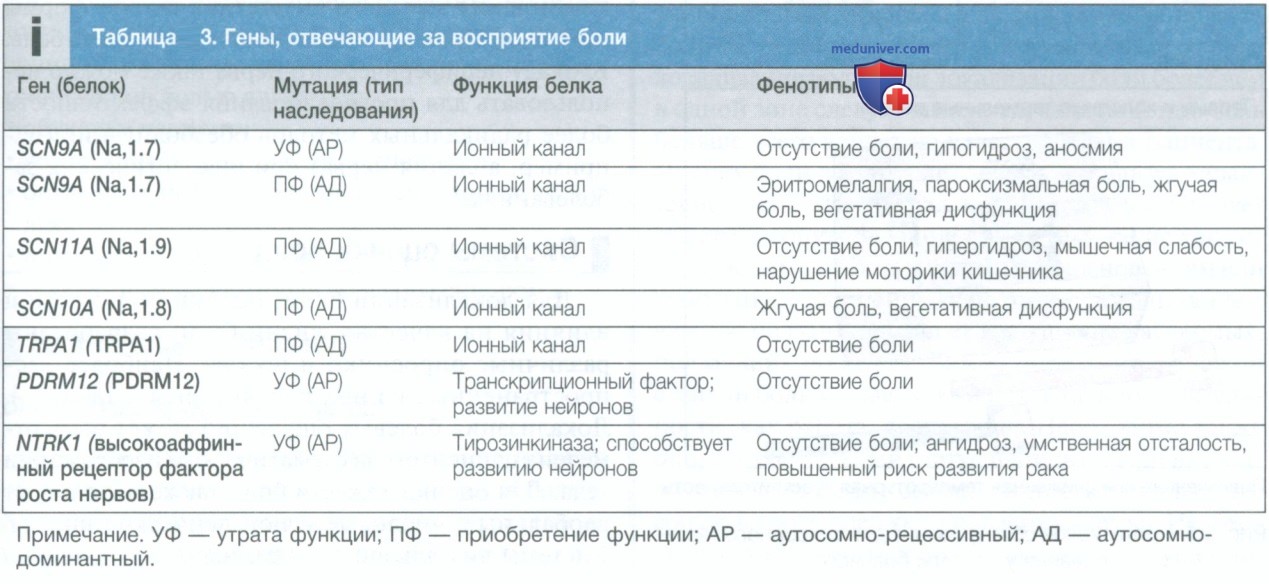
Было описано несколько редких синдромов, при которых нечувствительность к боли или усиление болевых реакций возникают в результате мутации одного гена, как показано в табл. 3. Большинство из них обусловлены мутациями, влияющими на работу ионных каналов, которые играют важную роль в нейротрансмиссии (см. рис. 3). Другие причины включают мутации в гене NTKR1, который кодирует рецептор фактора роста нервов, и мутации в факторе транскрипции PDRM12, участвующем в развитии нейронов.
Видео проводящие пути болевой и температурной чувствительности