Защиту от инфекционных агентов обеспечивает сочетание анатомических и физиологических барьеров: кожа, слизистые оболочки, эпителий ДП, мукоцилиарный клиренс, компоненты иммунной системы. Иммунная система позвоночных объединяет два механизма ответа. Врожденный (естественный) иммунитет срабатывает быстро: рецепторы закодированы генетически. Врожденная защита объединяет внутренние реакции клетки на вирусные инфекции, ответы лейкоцитов на патогены и растворимые медиаторы — белки комплемента.
Приобретенный (адаптивный) иммунитет специфичен для Т- и В-клеток. ДНК этих клеток подвергается рекомбинации, становится способной к генерации рецепторов. Для минимизации активности аутореактивных клеток необходим процесс обучения. Некоторые субпопуляции лимфоцитов по природе считаются врожденными. Они не требуют рекомбинации ДНК или ограничиваются единичным этапом рекомбинации для создания моно-специфического рецептора.
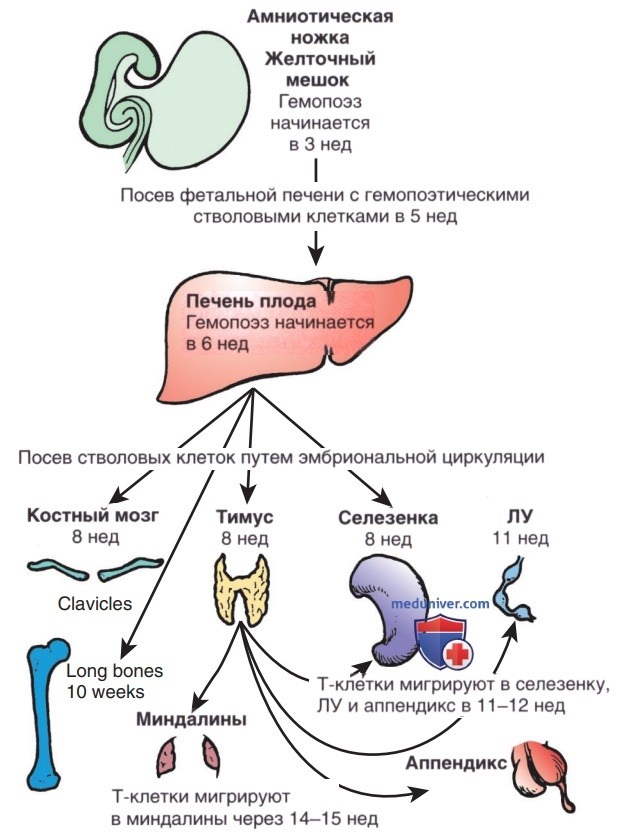
а) Лимфопоэз плода. Плюрипотентные гемопоэтические стволовые клетки сначала появляются в желточном мешке на 2,5-3 нед гестации, затем на 5-й нед гестации мигрируют в печень плода, позже обосновываются в костном мозге, где остаются на протяжении всей жизни (рис. 1). Лимфоидные стволовые клетки развиваются и дифференцируются в Т-, В-или NK-клетки (естественные киллеры), в зависимости от органов или тканей, к которым они транспортируются.
Рисунок 1. Схемы миграции гемопоэтических стволовых клеток и зрелых лимфоцитов во время в/утробного развития человека.
Развитие первичных лимфоидных органов (вилочковой железы и костного мозга) начинается в середине I триместра беременности и происходит быстро. Далее следует развитие вторичных лимфоидных органов: селезенки, ЛУ, миндалин, пейеровских бляшек и собственной пластинки. Эти органы — места дифференцировки Т-, В- и NK-лимфоцитов из стволовых клеток на протяжении всей жизни. Начальный органогенез и продолжающаяся клеточная дифференцировка — результат взаимодействия молекул клеточной поверхности лимфоцитов, молекул микроокружения и белков, секретируемых вовлеченными клетками. Кластеры дифференцировки (CD) — клеточные белки (табл. 1), а цитокины и хемокины — растворимые медиаторы иммунной функции (табл. 2).
1. Развитие и дифференцировка Т-клеток. Примитивный зачаток тимуса формируется из эктодермы 3-й жаберной щели и энтодермы 3-го жаберного кармана на 4 нед беременности. С 7-8 нед правый и левый зачатки сливаются по средней линии. Мигрировавшие из костного мозга в печень предшественники Т-клеток начинают колонизировать перитимическую мезенхиму на 8 нед беременности и перемещаются в тимус на 8-8,5 нед.
Клетки, которые быстрее всего попадают в тимус, находятся в субкапсулярной области и не экспрессируют ни CD3, CD4, CD8, ни какой-либо тип Т-клеточного рецептора (TCR). Предшественники лимфоидных клеток запрограммированы для пролиферации и превращения в тимоциты путем взаимодействия со стромой тимуса. Эта клеточная стадия длится до перестройки локуса β-цепи TCR. Затем β-цепь соединяется с суррогатной pre-Т-α-цепью.
На этом этапе проверяется функция β-цепи: если сигналы передаются, перестройка β-цепи прекращается. Затем CD4 и CD8 экспрессируются одновременно (дважды «+» тимоциты). Кортикальные тимоциты плода — самые быстро делящиеся клетки организма. Их количество увеличивается в 100 000 раз в течение 2 нед после попадания стволовых клеток в тимус. По мере размножения и созревания эти клетки мигрируют глубже в кору тимуса. Двойные «+» тимоциты начинают эффективную перестройку локуса a-цепи. Перестройка гена TCR происходит в процессе соединения больших несмежных блоков ДНК. Блоки V — variable, D — diversity и J — joining существуют в семействах минимально различных сегментов.
Случайные комбинации сегментов составляют большую часть огромного разнообразия TCR, которое позволяет распознавать миллионы различных АГн. Для реаранжировки гена TCR требуется присутствие генов, активирующих рекомбиназу, RAG1 и RAG2, и др. компонентов рекомбиназы.
Когда незрелые кортикальные тимоциты экспрессируют TCR, начинаются процессы «+» и «-» отбора. В незрелых тимоцитах происходит «+» отбор, который распознает АГн главного комплекса гистосовместимости (МНС, Major Histocompatibility Complex), присутствующие на кортикальных эпителиальных клетках тимуса. Некоторые клетки отбираются для созревания в одиночные «+» клетки CD4 или CD8. Затем в мозговом веществе тимуса на эпителиальных клетках происходит «-» отбор. Аутореактивные Т-клетки подвергаются апоптозу и умирают. Т-клетки мигрируют из вилочковой железы в селезенку, ЛУ и аппендикс на 11-12 нед эмбриональной жизни и в миндалины к 14-15 нед.
Они покидают вилочковую железу и через кровоток распределяются по всему телу, достигая наибольшей концентрации в паракортикальных областях ЛУ, периартериолярных областях селезенки и грудном лимфатическом протоке. Недавние эмигранты из тимуса коэкспрессируют изоформы CD45RA и CD62L (L-селектин).
Перестройка локуса TCR во время развития интра-тимических Т-клеток приводит к эксцизии ДНК, а иссеченные участки в качестве побочного продукта образуют кольцевые эписомы. Эти Т-рецепторные эксцизионные кольца выявляют в Т-клетках, недавно мигрировавших из тимуса на периферию (недавние тимусные эмигранты). Т-рецепторные эксцизионные кольца, обнаруженные в засохших пятнах крови, взятой у младенцев вскоре после рождения, — тест для скрининга новорожденных на тяжелый комбинированный иммунодефицит. К 12-й нед беременности Т-клетки могут пролиферировать в ответ на лектины растений — фитогемагглютинин и конканавалин А. АГн-специфические Т-клетки обнаруживаются до 20-й нед беременности.
Тельца Хассалла — завитки терминально дифференцированных медуллярных эпителиальных клеток. Они впервые обнаруживаются в мозговом веществе тимуса на 16-18 нед эмбриональной жизни.
2. Развитие и дифференцировка В-клеток. Развитие В-клеток в печени плода начинается на 7-й нед беременности. На 8-й нед в/утробной жизни стволовые клетки CD34 из печени плода заселяют костный мозг ключиц, а на 10-й нед появляются в костном мозге длинных костей (рис. 1). По мере дифференцировки В-клеток из примитивных стволовых клеток они проходят стадии, отличающиеся последовательной перестройкой генных сегментов Ig и образованием разнообразных АГн-рецепторов. Ранняя про-В-клетка — первый потомок плюрипотентных стволовых клеток, предназначенных для формирования В-линии. На этой стадии впервые перестраивается локус тяжелой цепи. В ранних про-В-клетках перестройки D-J происходят на обеих хромосомах.
В поздних про-В-клетках сегмент V перестраивается в сегмент гена D-J. Следующий этап дифференцировки — пре-В-клетка. В это время происходит перестройка генов легкой цепи Ig. Пре-В-клетка отличается экспрессией цитоплазматических μ тяжелых цепей, но без поверхностного IgM (sIgM), потому что легкие цепи Ig еще не продуцируются. Далее следует стадия незрелых В-клеток, когда гены легких цепей уже перегруппированы и вместо sIgD экспрессируется sIgM. Незрелые В-клетки покидают костный мозг, следуя к вторичным лимфоидным органам. Последняя стадия развития АГн-независимых В-клеток — зрелая наивная В-клетка, которая коэкспрессирует sIgM и sIgD. Пре-В-клетки обнаруживают в печени плода на 7 нед беременности, sIgM+ и sIgG+ В-клетки — на 7-11 нед, sIgD+ и sIgA+ В-клетки — на 12-13 нед.
К 14 нед эмбриональной жизни процент циркулирующих лимфоцитов, несущих sIgM и sIgD, аналогичен пуповинной крови и немного выше, чем в крови взрослых.
АГн-зависимые стадии развития В-клеток — стадии, регистрируемые после стимуляции АГн-зрелых В-клеток во вторичных лимфоидных органах. После активации АГн-зрелые В-клетки могут стать В-клетками памяти или плазмобластами. Оба процесса происходят при участии Т-хелперов.
По типу тяжелой цепи различают 5 классов (изотипов) Ig: IgM, IgG, IgA, IgD и IgE. IgG и IgM — единственные связывающие комплемент изотипы, наиболее важные Ig в крови и др. внутренних жидкостях организма. Они защищают организм от инфекционных агентов. IgM из-за своего большого размера присутствует во в/сосудистом русле, a IgG обнаруживается во всех внутренних жидкостях тканей организма. IgA — основной защитный Ig внешней секреции в ЖКТ, респираторном и урогенитальном трактах, содержится в кровотоке. IgE содержится во внутренних и внешних тканевых жидкостях организма, играет важную роль в защите хозяина от паразитов. Из-за высокого сродства рецепторов IgE к базофилам и тучным клеткам IgE — основной медиатор аллергических реакций немедленного типа.
Предназначение IgD до сих пор окончательно не ясно. IgG подразделяются на 4 подкласса (IgG1, IgG2, IgG3 и IgG4), a IgA — на два (IgAl и IgA2). Каждый из этих подклассов выполняет разные биологические функции. Секретируемые IgM и IgE обнаруживают на 10 нед беременности, a IgG — в период между 11 и 12 нед.
Стадии развития В-клеток описаны в контексте онтогенеза В-клеток в утробе матери, но процесс развития В-клеток из плюрипотентных стволовых клеток продолжается на протяжении всей постнатальной жизни. Плазматические клетки не обнаруживаются в лимфоидных тканях плода до 20 нед беременности. Позднее их присутствие редко наблюдается из-за стерильной среды матки. Лимфоидное развитие кишечника происходит относительно поздно. Пятна Пейера обнаружены в значительном количестве к 5 мес в/утробного развития, а плазматические клетки обнаружены в собственной пластинке к 25 нед беременности. До рождения в ЛУ могут находиться первичные фолликулы, а вторичные фолликулы отсутствуют.
Плод человека получает значительные количества материнского IgG на 12 нед беременности трансплацентарно. Это количество постоянно увеличивается, пока при рождении сыворотка пуповинной крови не будет содержать концентрацию IgG, сравнимую с концентрацией материнской сыворотки или превышающую ее. IgG — единственный класс АТл, который способен проникать через плаценту в достаточном количестве. Все 4 подкласса IgG проникают через плаценту, но у IgG2 эта способность ограничена. Небольшое количество IgM (10% от уровня взрослого) и несколько нанограммов IgA, IgD и IgE обнаруживают в сыворотке пуповинной крови. Поскольку ни один из этих белков не проникает через плаценту, предполагается их фетальное происхождение.
Эти наблюдения позволяют предположить, что определенные АГн-стимулы в норме проникают через плаценту, вызывая реакции даже у неинфицированных плодов. У некоторых младенцев с атопией отмечены АТл IgE к АГн яичного белка, воздействию которого они не подвергались в послеродовой период. Вероятно, синтез АТл у плода индуцирован АГн, попавшими в организм матери.
3. Развитие естественных киллеров. Активность NK-клеток в клетках печени плода обнаруживают в период между 8 и 11 нед беременности. NK-лимфоциты также развиваются из клеток-предшественников костного мозга. Процессинг тимуса не считается необходимым условием для развития NK-клеток, хотя NK-клетки обнаруживаются в тимусе. После выхода из костного мозга NK-клетки попадают в кровоток или мигрируют в селезенку, при этом в ЛУ обнаруживается мало NK-клеток. У здоровых людей NK-клетки составляют 8-10% всех лимфоцитов. Некоторые ткани содержат большое количество NK-клеток. В отличие от Т- и В-клеток, NK-клетки не перестраивают гены АГн-рецепторов во время своего развития, а определяются их функциональной способностью опосредовать не-АГн-специфическую цитотоксичность.
NK-клетки содержат рецепторы, ингибирующие киллеры, которые распознают определенные АГн главного комплекса гистосовместимости и ингибируют уничтожение собственных тканей.
NK-активирующие рецепторы распознают стрессовый белок, а баланс взаимодействия активирующих и ингибирующих рецепторов определяет реакции NK-клеток. Если вирусная инфекция снижает экспрессию главного комплекса гистосовместимости класса I, потеря ингибиторной функции приводит к цитотоксичности. Высокий уровень стрессовых белков, наблюдаемый при вирусных инфекциях, также может активировать цитотоксичность.
4. Особенности движения лимфоцитов. Основные функции Т-лимфоцитов — стимуляция В-клеток для выработки АТл, уничтожения инфицированных вирусом или опухолевых клеток и активация макрофагов, предназначенных для в/клеточного уничтожения. Субпопуляции регуляторных Т-клеток (Tregs) значима для предотвращения аутоиммунных реакций. Т-клетки активируются АГн, представленным АГн-презентирующими клетками. Это дендритные клетки, макрофаги или В-клетки. Для высокоаффинного связывания Т-клеток с АГн-презентирующими клетками несколько молекул на Т-клетках, помимо TCR, связываются с молекулами на клетках-мишенях. Молекула CD4 связывается с молекулами главного комплекса гистосовместимости класса II на АГн-презентирующих клетках. CD8 на цитотоксических Т-клетках связывает молекулу главного комплекса гистосовместимости класса I на клетке-мишени.
Ассоциированный с функцией лимфоцитов АГн 1 (LFA-1) на Т-клетке связывает белок, называемый ICAM-1 (молекула в/клеточной адгезии 1), обозначенный как CD54, на АГн-презентирующих клетках. CD2 на Т-клетках связывает LFA-3 (CD58) на АГн-презентирующих клетках. Благодаря адгезии Т-клеток с АГн-презентирующими клетками (иммунологический синапс), активируются Т-хелперные (Th) клетки на синтез IL. Также активируются молекулы клеточной поверхности — лиганды CD40 (CD 154), которые помогают В-клеткам, а цитотоксические Т-клетки активируются на уничтожение своих мишеней. Ключевая защитная сеть для соответствующей активации Т-клеток в условиях реальной угрозы — требование ко-стимуляции Т-клеток. АГн-презентирующие клетки, которые столкнулись с патогеном, экспрессируют CD80 и CD86.
Вовлечение этих молекул дает второй костимулирующий сигнал. Без ко-стимуляции Т-лимфоциты станут энергическими или нефункциональными.
При первичном ответе АТл нативный АГн переносится в ЛУ, дренирующий этот участок, связывается комплементом, фагоцитируются специализированными клетками, называемыми фолликулярными дендритными клетками, и экспрессируется на их поверхности. Зрелые В-клетки, несущие sIgM, специфичные для этого АГн, затем связываются с АГн на поверхности фолликулярных дендритных клеток. Если сродство В-клеточного sIgM-АТл к АГн, присутствующему на фолликулярных дендритных клетках, достаточно, и активированные Т-клетки передают др. сигналы, В-клетка трансформируется в В-клетки памяти или продуцирующие АТл плазматические клетки. Сигналы при активации Т-клеток — секретируемые ими цитокины: IL-4, IL-5, IL-6, IL-10, IL-13 и IL-21 (табл. 2), а также поверхностные молекулы Т-клеток, лиганды CD40 или CD154. При контакте активированной Т-клетки CD4+ с В-клеткой они связываются с рецептором CD40 на поверхности В-клетки.
Взаимодействие CD40 на В-клетках с CD154 на Т-клетках в присутствии специфических цитокинов стимулирует В-клетки к пролиферации и к синтезу Ig. При первичном иммунном ответе вырабатываются только АТл IgM, большая часть которых имеет относительно низкое сродство к АГн. Некоторые В-клетки становятся В-клетками памяти во время первичного иммунного ответа. Вторичный ответ АТл возникает, когда В-клетки памяти снова встречаются с данным АГн. Развивающиеся В-клетки памяти переключают свои гены Ig так, что при вторичном воздействии того же АГн начинается выработка АТл IgG, IgA или IgE с более высокой аффинностью. Как и при первичном ответе, формируются плазматические клетки. Быстро образуется большое количество клеток и вырабатываются АТл IgG, IgA и IgE. Изменения генов Ig (соматическая гипермутация) приводят к увеличению аффинности этих АТл.
Истинная картина изотипического ответа на АГн у нормальных людей варьирует в зависимости от типа АГн и цитокинов, присутствующих в микроокружении. Переключение класса и соматическая гипермутация зависят от участия Т-клеток. Т-клетки — «стражи» для выработки специфических АТл.
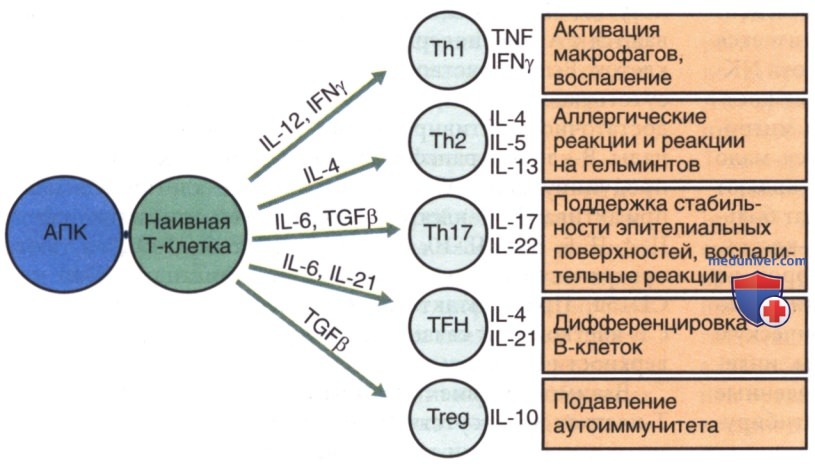
б) Особенности лимфоцитов новорожденного. Практически все Т-клетки плацентарной крови содержат CD45RA (наивную) изоформу, и преобладание CD45RA над Т-клетками CD45RO сохраняется в детстве. По достижении среднего возраста у человека преобладают Т-клетки CD45RO (памяти). CD4 Т-клетки далее подразделяются в соответствии с производимыми при активации цитокинами. Клетки ТЫ продуцируют IL-2 и интерферон (IFN)-γ, которые способствуют развитию цитотоксических Т-лимфоцитов или реакций ГЗТ. Клетки Th2 продуцируют IL-4, IL-5, IL-6, IL-13, и IL-21 (табл. 2), которые обуславливают ответы В-клеток с развитием атопии и аллергической сенсибилизации. Клетки Th17 продуцируют IL-17 и Tregs, синтезируя IL-10 (рис. 2). Дифференциацию подклассов клеток памяти определяет среда цитокинов, регулирующая специфические факторы транскрипции и эпигенетические изменения.
Рисунок 2. Дифференцировка Т-клеток в субпопуляцию Т-клеток памяти строго регулируется цитокинами. Специфические субпопуляции играют разные роли в защите хозяина. АПК — АГн-представляющая клетка; IFN — интерферон; IL — интерлейкин; Th — Т-хелперная клетка; TFH — фолликулярная хелперная Т-клетка; TGF — трансформирующий фактор роста; TNF — фактор некроза опухоли
Эти подклассы в стабильны in vivo, но в некоторых случаях могут переходить в др. подклассы. Клетки памяти быстрее реагируют на АГн и ориентированы на выработку цитокинов, способствующих элиминации патогенов.
Для новорожденных характерна повышенная восприимчивость к инфекциям, вызванным гр/о-микроорганизмами, поскольку АТл IgM, мощные опсонины, усиливающие фагоцитоз, не проникают через плаценту. Др. основной опсонин, C3b, в сыворотке новорожденных более низкий, чем у взрослых. Эти факторы объясняют нарушение фагоцитоза некоторых микроорганизмов новообразованными полиморфно-ядерными клетками. АТл IgG, передающиеся от матери, подходят для нейтрализации большинства гр/п-бактерий, а АТл IgG к вирусам обеспечивают защиту от этих агентов. Поскольку в младенчестве наблюдается относительный дефицит подкласса IgG2, АТл к капсульным полисахаридным АГн может быть недостаточно. Поскольку недоношенные дети к моменту рождения получали меньше материнского IgG, чем доношенные дети, их опсоническая активность сыворотки крови низка для всех типов микроорганизмов.
Вскоре после рождения дети синтезируют АТл класса IgM с повышенной скоростью в ответ на огромную АГн-стимуляцию нового окружения. Разницы в выработке IgM у доношенных и недоношенных младенцев не отмечено. Через 6 сут после рождения концентрация IgM в сыворотке крови резко возрастает. Это повышение продолжается до достижения значений взрослого человека (к ~1 году). Плацентарная сыворотка здоровых неинфицированных новорожденных не содержит обнаруживаемого уровня IgA. Сывороточный уровень IgA впервые выявляется на 13 сут постнатальной жизни, но остается низким в младенчестве. Плацентарная сыворотка содержит IgG в концентрации, сравнимой или большей, чем в материнской сыворотке.
Материнский IgG постепенно снижается в первые 6-8 мес жизни, а скорость синтеза детских IgG увеличивается (IgG1 и IgG3 быстрее, чем IgG2 и IgG4 в течение первого года), пока концентрации общего IgG взрослых не будут достигнуты и сохранятся к 7-8 годам. Первыми достигают значений уровня взрослого человека IgG1 и IgG4, через 10 лет — IgG3, через 12 лет — IgG2. Уровень IgG в сыворотке у младенцев достигает нижнего значения через 3-4 мес постнатальной жизни. Скорость нарастания IgE соответствует скорости увеличения IgA.
После достижения взрослой концентрации каждого из 3 основных Ig эти уровни остаются постоянными у здорового человека. Способность продуцировать специфические АТл к белковым АГн сохраняется при рождении, но младенцы не могут вырабатывать АТл к полисахаридным АГн до 2 лет, если полисахарид не конъюгирован с белком-носителем, как в случае конъюгата Haemophilus influenzae типа b и вакцины против Streptococcus pneumoniae.
В плацентарной крови процент NK-клеток ниже, чем в крови детей и взрослых, но из-за более высокого количества лимфоцитов абсолютное количество NK-клеток примерно одинаково. Способность NK-клеток пуповинной крови опосредовать лизис мишеней в анализах NK-клеток или АТл-зависимых клеточных реакциях цитотоксичности составляет 2/3 от способности взрослых.
1. Развитие лимфоидных органов. Пропорция лимфоидной ткани в организме относительно мала. Лимфоидная ткань хорошо развита при рождении и быстро созревает в послеродовой период. Тимус имеет самые большие размеры по отношению к размеру тела плода, при рождении составляет 2/3 его зрелого веса, достигаемого в первый год жизни. Максимального объема тимус достигает незадолго до полового созревания, а затем наступает его инволюция. К первому году жизни все лимфоидные структуры становятся гистологически зрелыми. Абсолютное количество лимфоцитов в периферической крови достигает максимального значения в течение первого года жизни (рис. 2).
Селезенка постепенно набирает массу во время созревания, достигая максимума только у взрослых. Среднее количество пейеровских пятен при рождении составляет 1/2 от такового количества у взрослого и постепенно увеличивается, пока в подростковом возрасте не превысит среднее число для взрослых.
в) Наследование аномалий в развитии Т-, В- и NK-клеток. Описано >300 синдромов иммунодефицита. Специфические молекулярные дефекты выявлены для большинства заболеваний. Большинство из них— Х-сцепленные рецессивные патологии, также встречаются АуД-заболевания с потерей функции и АуД-болезни с усилением функции. Недостаточности ассоциируются с отсутствием типа или линии клеток (отсутствие Т-клеток при тяжелом комбинированном иммунодефиците), отсутствием субклассов клеток (отсутствие Treg при иммунной дисрегуляции, полиэндокринопатии, Х-сцепленном синдроме) или дисфункцией клеток (гемофагоцитарный лимфогистиоцитоз). В некоторых случаях поражены несколько типов клеток, а при отдельных синдромах избыток клеток определенного типа или функции нарушает предельный баланс, необходимый для иммунного гомеостаза.