а) Транслокации. Транслокации, заключающиеся в переносе материала от одной хромосомы к другой, происходят с частотой 1:500 живорожденных детей. Они м.б. унаследованы от родителя-носителя / возникнуть de novo, оставляя незатронутыми др. членов семьи. Транслокации подразделяются на два типа: реципрокные и робертсоновские, с участием двух хромосом (рис. 1).
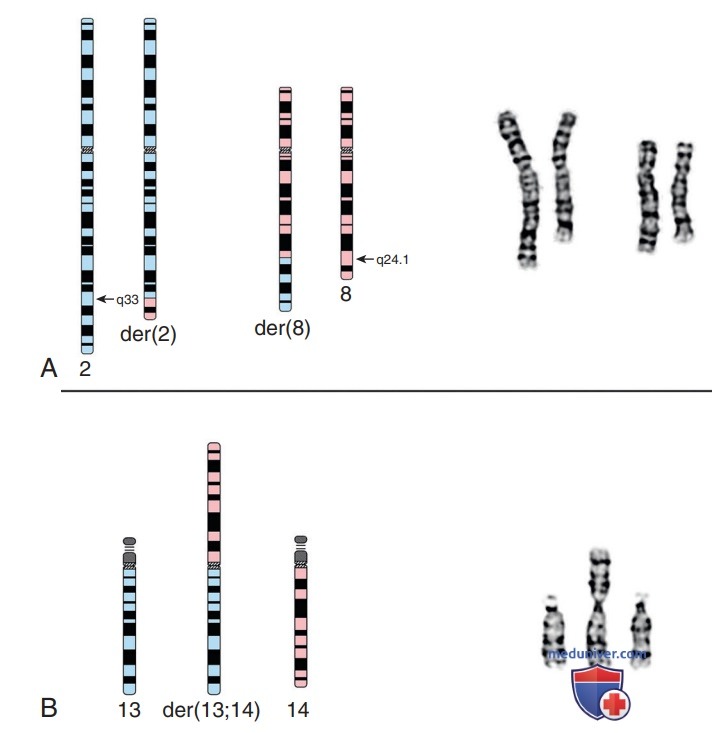
Рисунок 1. А — схематическая диаграмма (слева) и частичный G-окрашенный кариотип (справа) реципрокной транслокации между хромосомой 2 (синий) и хромосомой 8 (розовый). Точки разрыва находятся на длинном (q) плече обеих хромосом на полосах 2q33 и 8q24.1, с реципрокным обменом материалом между дериватными (der) хромосомами 2 и 8. Это перемещение сбалансировано, без прибавки/потери материала. Номенклатура этого обмена — t(2;8)(q33;q24.1); В — схематическая диаграмма (слева) и частичный G-окрашенный кариотип (справа) робертсоновской транслокации между хромосомами 13 (синий) и 14 (розовый). Точки разрыва находятся на центромере (полоса q10) обеих хромосом, со слиянием длинных плеч в одну производную хромосому и потерей материала коротких (р) плеч. Номенклатура этого обмена: der( 13;14)(q 10; q10).
Реципрокные транслокации являются результатом разрывов в негомологичных хромосомах с реципрокным обменом оторванными сегментами. Носители реципрокной транслокации обычно фенотипически нормальны, но подвергаются повышенному риску выкидыша, вызванного передачей несбалансированных реципрокных транслокаций, и вынашивания потомства с хромосомными аномалиями. Несбалансированные транслокации являются результатом аномальной сегрегации / кроссин-говера в половых клетках носителей хромосомной транслокации.
Робертсоновские транслокации затрагивают две акроцентрические хромосомы (хромосомы 13, 14, 15, 21 и 22), которые сливаются в центромерной области с последующей потерей коротких плеч. Поскольку короткие плечи всех пяти пар акроцентрических хромосом имеют несколько копий генов, кодирующих рибосомную РНК, потеря короткого плеча двух акроцентрических хромосом не оказывает повреждающего воздействия. Результирующий кариотип имеет всего 45 хромосом, включая транслоцированную хромосому, которая состоит из длинных плеч двух слитых хромосом.
Носители робертсоновских транслокаций обычно фенотипически нормальны. Однако они подвержены повышенному риску невынашивания беременности и несбалансированных хромосомных перестроек у фенотипически ненормального потомства.
В некоторых редких случаях транслокации могут включать три хромосомы и более, так называемые комплексные перестройки. Другой, менее распространенный тип — инсерционная транслокация (инсерция). Инсерции возникают в результате вырезания фрагмента хромосомного материала, который затем снова вставляется в ту же хромосому в другом месте / в другую хромосому.
б) Инверсии. При инверсии одна хромосома разрывается в двух точках; затем оторванный участок переворачивается и присоединяется к той же хромосоме. Инверсия происходит у 1:100 живорожденных младенцев. Существует два типа инверсий: перицентрические и парацентрические. При перицентрических инверсиях разрывы находятся в двух противоположных плечах хромосомы и захватывают центромеру. Обычно их обнаруживают, потому что они меняют положение центромеры. Разрывы при парацентрических инверсиях происходят только в одном плече. Носители инверсий обычно фенотипически нормальны, но они подвержены повышенному риску невынашивания, обычно при парацентрических инверсиях, и рождения потомков с хромосомными аномалиями при перицентрических инверсиях.
в) Делеции и дупликации. Делеции подразумевают потерю хромосомного материала. В зависимости от местонахождения они м.б. классифицированы как терминальные (в конце хромосомы) / интерстициальные (в пределах плеча хромосомы). Делеции бывают изолированными / могут возникать совместно с дупликацией другого сегмента хромосомы. Последнее обычно происходит при несбалансированной реципрокной хромосомной транслокации, вторичной по отношению к транслокации/инверсии у носителя и возникшей в результате аномального кроссинговера/сегрегации.
Носитель делеции является моносомным для генетической информации отсутствующего сегмента. Делеции обычно связаны с умственной отсталостью и пороками развития. Наиболее часто наблюдаемыми в обычных препаратах хромосом делециями являются 1р-, 4р-, 5р-, 9р-, 11p-, 13q-, 18р-, 18q- и 21q- (табл. 12, рис. 2), а также все дистальные/терминальные делеции коротких/длинных плеч хромосом. Делеции могут наблюдаться в обычных препаратах хромосом, а делеции и транслокации размером >5-10 Мб обычно видны под микроскопом.
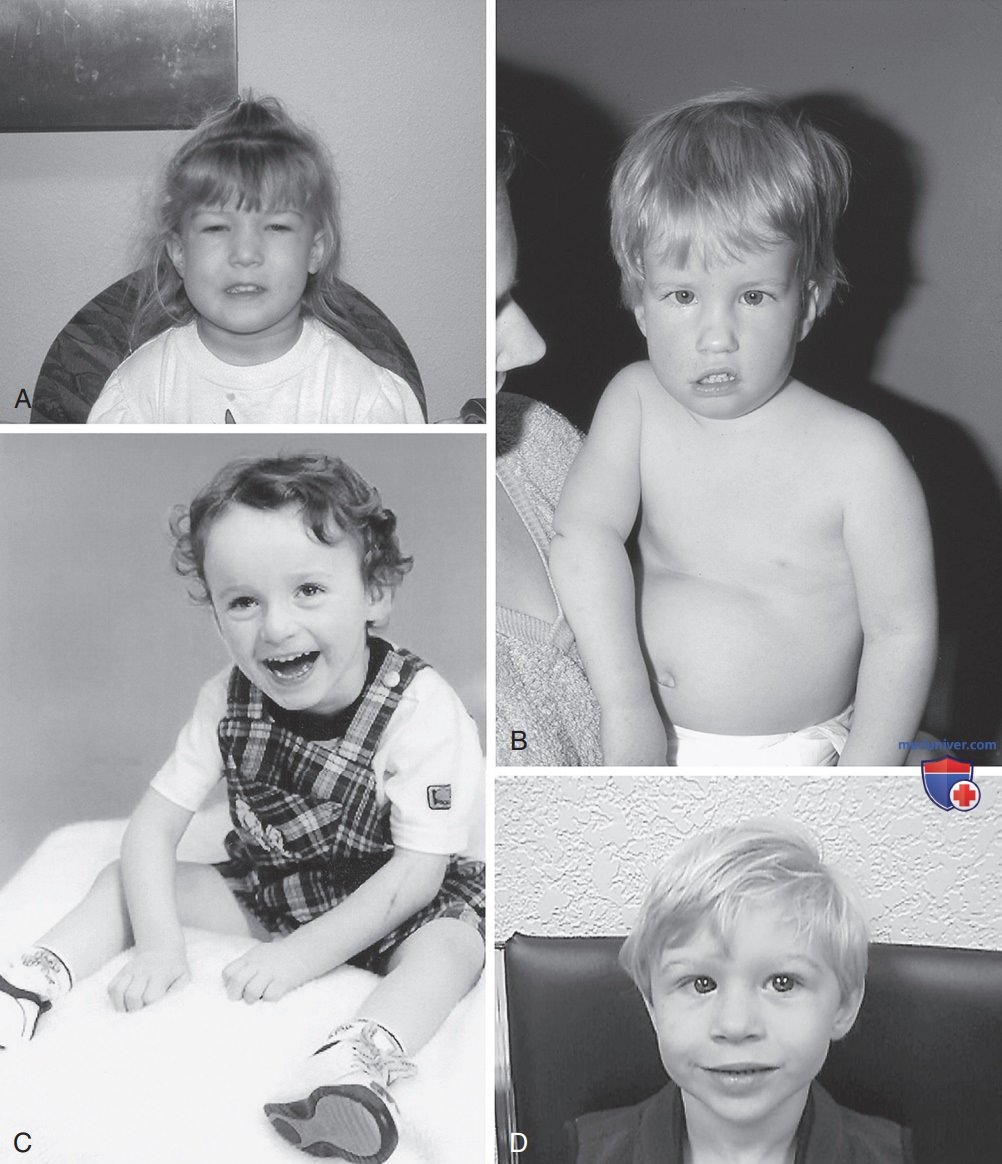
Рисунок 2. А — ребенок с велокардиофациальным синдромом (делеция 22q11.2); В — ребенок с синдромом Прадера-Вилли (делеция 15q 11-13); С — ребенок с синдромом Ангельмана (делеция 15q11-13); D — ребенок с синдромом Вильямса (делеция 7q11.23).
Методы дифференциального окрашивания хромосом с высоким разрешением, FISH и молекулярные методы исследования, такие как aCGH, могут выявить делеции, слишком маленькие для выявления на обычных/рутинных препаратах хромосом (см. рис. ниже). Микроделеции влекут за собой потерю небольших участков хромосомы, самые большие из которых можно обнаружить только с помощью анализа профазных хромосом и молекулярных методов. В случае субмикроскопических делеций недостающий фрагмент можно обнаружить только с помощью молекулярных методологий, таких как исследования на основе ДНК (напр., aCGH, FISH). Наличие дополнительного генетического материала из той же хромосомы называется дупликацией. Дупликации также м.б. спорадическими / возникать в результате патологической сегрегации у носителей транслокации/инверсии.
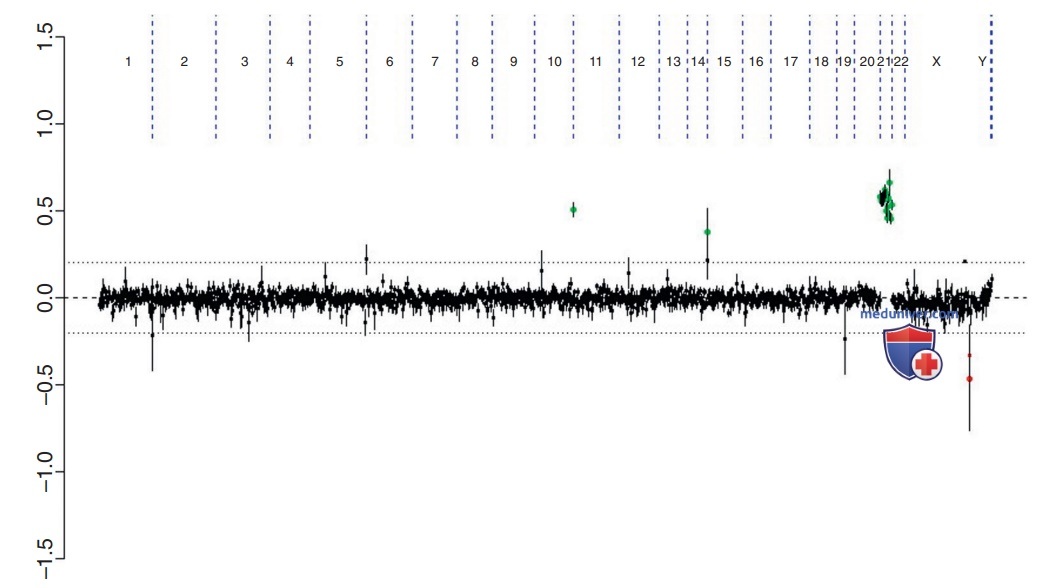
aCGH у пациентки с синдромом Дауна. Каждая черная точка представляет собой часть сегмента ДНК, специфичную для разного расположения хромосомы. Считается, что большинство точек, отображаемых между осями 0,0 и 0,2, находятся в пределах нормы. Исключения часто являются результатом полиморфных вариаций. Группа зеленых точек в кластерах на хромосоме 21 и выше 0,5. Они представляют собой увеличение числа копий сегментов ДНК для хромосомы 21, наблюдаемое при синдроме Дауна, соответствующее трисомии 21
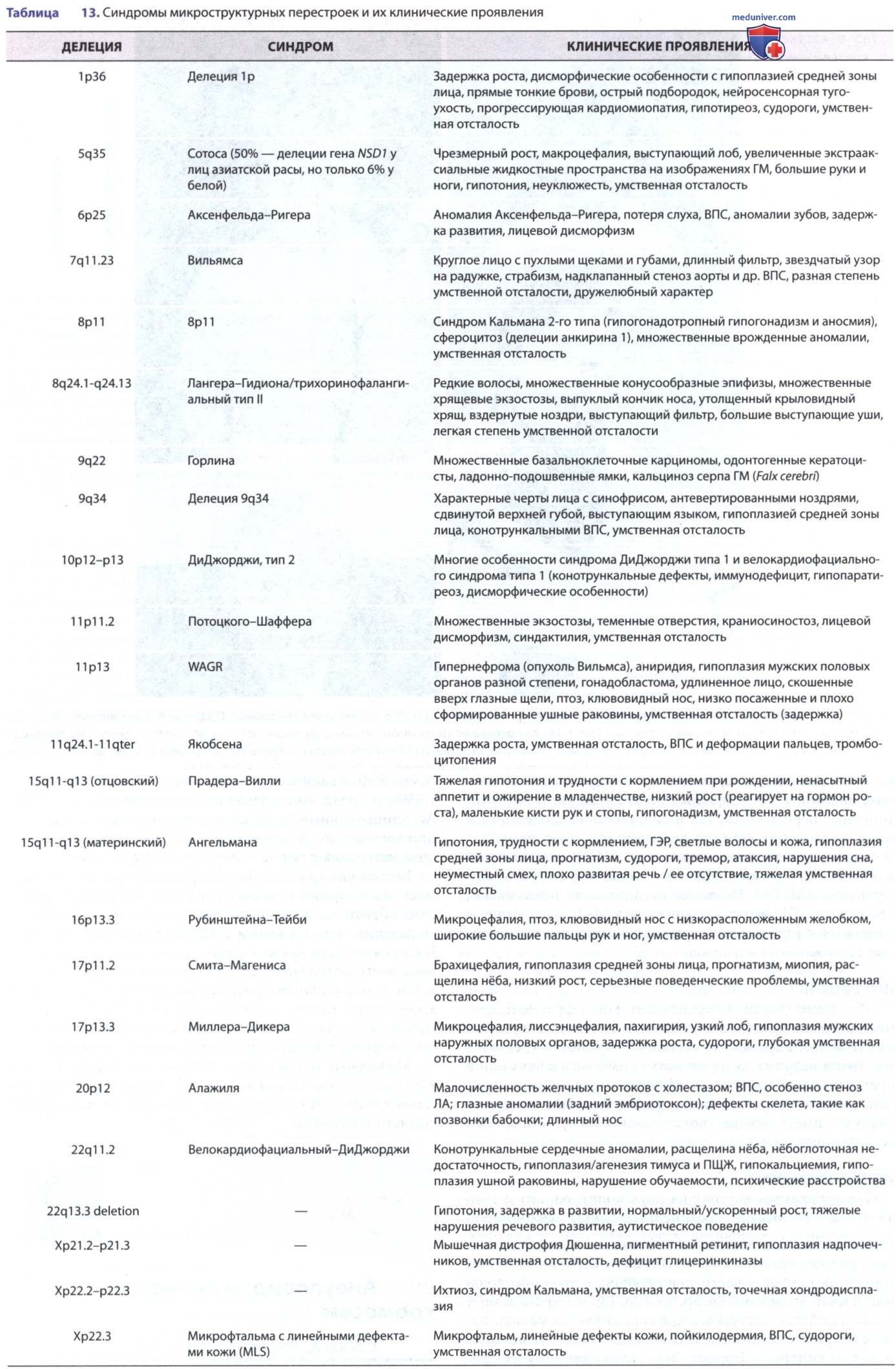
Микроделеции и микродупликации обычно затрагивают области, включающие несколько генов, поэтому больные люди могут иметь особый фенотип в зависимости от количества задействованных генов. Когда такая делеция затрагивает более одного гена, состояние называется синдромом микроструктурных перестроек (табл. 13). После того как в клинической практике стал доступен метод aCGH, было обнаружено большое количество дупликаций, большинство из которых являются микродупликациями. Многие из этих синдромов микродупликаций являются реципрокными дупликациями известных делеций/аналогов микроделеций и имеют отличительные клинические признаки (табл. 14).
Субтеломерные участки часто вовлечены в хромосомные перестройки, которые невозможно визуализировать с помощью рутинной цитогенетики. Теломеры, являющиеся дистальными концами хромосом, представляют собой области, богатые генами. Дистальная структура теломер является практически одинаковой для всех хромосом, но проксимально к ним расположены уникальные области, известные как субтеломеры, которые обычно участвуют в делециях и др. хромосомных перестройках. Небольшие субтеломерные делеции, дупликации/перестройки (транслокации, инверсии) м.б. сравнительно обычным явлением у детей с неспецифической умственной отсталостью и малыми патологиями.
С помощью метода aCGH субтеломерные перестройки были обнаружены у 3-7% детей с умственной отсталостью средней и тяжелой степени и у 0,5% детей с легкой умственной отсталостью.
Мутации и аномалии длины теломер также вызывают врожденный дискератоз и др. синдромы апластической анемии, а также фиброз легких/печени. Как субтеломерные перестройки, так и синдромы микроделеции и микродупликации обычно диагностируются с помощью молекулярных методов, таких как aCGH и технологии мультиплексной амплификации зонда, зависящей от лигирования (MLPA). Недавние исследования показывают, что метод aCGH может обнаруживать 14-18% аномалий у пациентов с нормальными результатами ранее проведенных хромосомных анализов.
г) Инсерции. Инсерции (вставки) происходят, когда фрагмент хромосомы, разорванный в двух точках, включается в разрыв в другой части хромосомы. В общей сложности требуется три точки разрыва, которые могут возникать в двух или в пределах одной хромосомы. Инсерции типа нереципрокной транслокации встречаются редко. Носители инсерции рискуют иметь потомство с делециями/дупликациями вставленного сегмента.
д) Изохромосомы. Изохромосомы состоят из двух копий одного и того же плеча хромосомы, соединенных одной центромерой и являющихся взаимным зеркальным отображением. Наиболее часто наблюдаемые аутосомные изохромосомы, как правило, связаны с хромосомами с короткими плечами. Среди наиболее часто участвующих в этом образовании плечей хромосом: 5р, 8р, 9р, 12р, 18р и 18q. Выделяют также наиболее частую изохромосомную аномалию, наблюдаемую в длинном плече Х-хромосомы и вызывающую развитие синдрома Тернера. Лица, имеющие одну изохромосому X в 46 хромосомах, являются моносомными для генов в потерянном коротком плече и трисомными для генов, присутствующих в длинном плече Х-хромосомы.
е) Маркерные и кольцевые хромосомы. Маркерные хромосомы встречаются редко и обычно представляют собой хромосомные фрагменты, которые слишком малы для того, чтобы их можно было идентифицировать с помощью стандартной цитогенетики; обычно они встречаются как дополнительные к нормальным 46 хромосомам. Большинство из них носит спорадический характер (70%); часты случаи мозаицизма (50%) из-за митотической нестабильности маркерной хромосомы. Заболеваемость среди новорожденных составляет 1:3300, а среди лиц с умственной отсталостью — 1:300. Ассоциированный фенотип варьирует от нормального до патологического, в зависимости от количества хромосомного материала и генов, включенных во фрагмент.
Кольцевые хромосомы, обнаруживаемые во всех хромосомах человека, встречаются редко. Кольцевая хромосома образуется, когда оба конца хромосомы подвергаются делеции, а затем концы соединяются, образуя кольцо. В зависимости от нехватки/избыточности количества хромосомного материала (если кольцо присутствует в дополнение к нормальным хромосомам) пациент с кольцевой хромосомой может казаться нормальным / почти нормальным, а также может страдать умственной отсталостью и иметь множественные врожденные аномалии.
Маркерные и кольцевые хромосомы можно найти в клетках плотных опухолей у детей, клетки здоровых органов которых не содержат этого дополнительного хромосомного материала.