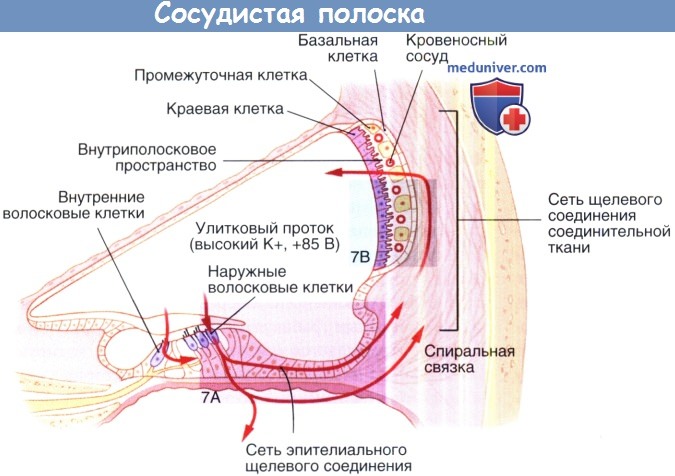
Сосудистая полоска играет ключевую роль в гомеостазе улитки, генерируя внутриулитковый потенциал и сохраняя специфический ионный состав эндолимфы. Сосудистая полоска является высоко васкуляризированной многослойной тканью, входящей в состав латеральной стенки улиткового протока. Она включает три различных типа клеток (краевые, промежуточные и базальные), каждый из которых необходим для ее функционирования.
Щелевые соединения обеспечивают ионный барьер, который отделяет сосудистую полоску, одни находятся на уровне маргинальных клеток, а другие на уровне базальных клеток. Внеклеточное пространство между этими двумя барьерами называется интрастриарным (внутриполосковым) пространством. Как показано на рисунке ниже, маргинальные клетки отделяют улитковый проток, наполненный эндолимфой, от интрастриарного пространства, которое наполнено интрастриарной жидкостью, в то время как базальные клетки отделяют интрастриарное пространство от перелимфы, окружающей фиброциты спиральной связки.
Промежуточные клетки, так же как и кровеносные сосуды входят в состав интрастриарного пространства. Щелевые межклеточные промежутки соединяют базальные клетки с промежуточными клетками и с фиброцитами спиральной связки, обеспечивая электрическую передачу, а также обмен ионов и небольших молекул. Система регуляции гомеостаза кохлеарной жидкости также включает эндолимфатический мешок, который отвечает за изменения объема эндолимфы и, возможно, изменяет гомеостаз при нарушении его функционирования.
Нарушения гомеостаза улитковой жидкости, вызванные изменениями внутриулиткового потенциала, ионного состава или механизмов его регуляции, ведут к различным формам тугоухости у людей и животных.
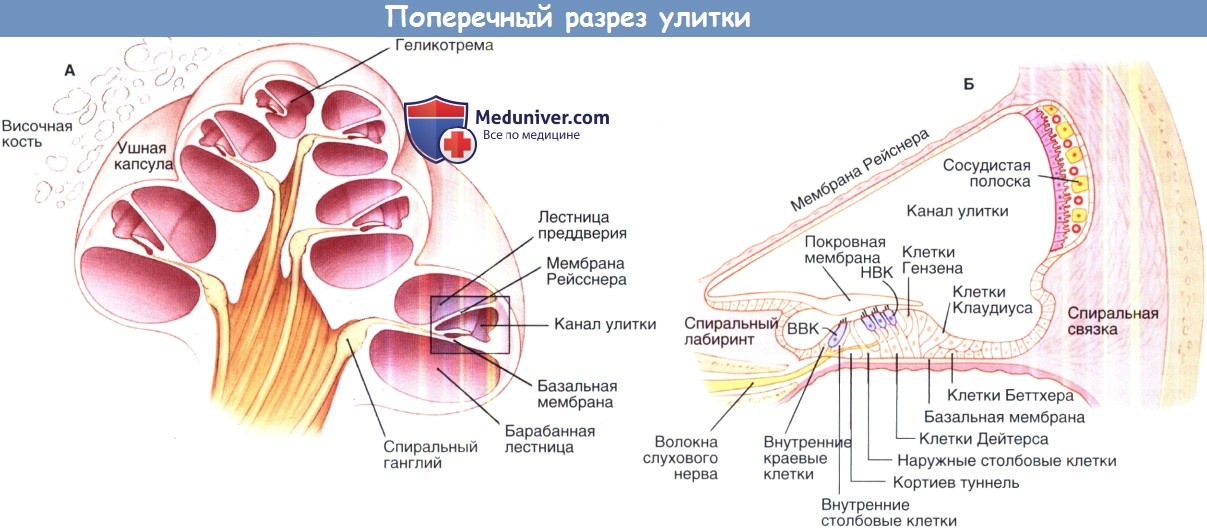
Поперечный разрез улитки. А, схематичное изображение улитки в разрезе.
Выделенная область, показанная на рисунке Б, изображает улитковый проток и окружающие структуры, такие как кортиев орган и сосудистую полоску в разрезе.
ВВК — внутренние волосковые клетки; НВК — наружные волосковые клетки.
а) Внутриулитковый потенциал и гомеостаз калия. Эффективная механоэлектрическая трансдукция (передача) волосковых клеток обеспечивается благодаря большой движущей силе катионов, входящих в цитоплазму клеток из улиткового протока (медиальной лестницы). Внутриулитковый потенциал эндолимфы, составляющий приблизительно +85мВт, и химический градиент К+ являются основными компонентами движущей силы, которая достигает 130 мВт, так как в покое потенциал волосковой клетки составляет -45 мВт. При снижении внутриулиткового потенциала на 1 мВт пороги слышимости увеличиваются примерно на 1 дБ.
К+, являющийся основным катионом эндолимфы, несет в себе большую часть электрического заряда, который генерирует внутриулитковый потенциал. Поэтому важно понимать, каким образом К+ движется через улитку. К+ может входить в волосковые клетки через каналы механоэлектрической передачи и высвобождаться через базолатеральные мембраны волосковых клеток в перелимфатическое внеклеточное пространство. Предполагается, что К+ может входить в поддерживающие клетки и двигаться по направлению к спиральной связке через широкие межклеточные пространства.
Кроме того, К+ может диффундировать внеклеточно через перелимфатическое пространство.
Фиброциты I и II типов спиральной связки захватывают К+ и обеспечивают его внутриклеточное перемещение в базальные и промежуточные клетки сосудистой полоски. К+ выпускается промежуточными клетками через KCNJ10 каналы в интрастриарное пространство, из которого он активно засасывается и ко-транспортируется в маргинальные клетки. Маргинальные клетки выделяют К+ в улитковый проток. В целом циркуляция К+ на самом деле не является истинной циркуляцией, так как перелимфа и интрастриарное пространство не образуют закрытую петлю, поскольку эти отсеки связаны с другими внеклеточными пространствами и с кровотоком.
Сбои в работе некоторых К+ каналов ведут к нарушению улиткового К+ гомеостаза, приводя к ухудшению слуха. У мышей потеря гена KCNE1, который кодирует субъединицу К+ канала, экспрессируемую маргинальными клетками, формирует очень схожий с кардиоаудиторным синдромом фенотип, характеризующийся снижением слуха и аритмией. Это наблюдение привело к выявлению двух генов человека, KNCE1 и KCNQ1, мутации которых вызывают кардиоаудиторныйсиндром.Вполне возможно, что KCNQ1 и KNCE1 формируют каналы, которые обеспечивают секрецию калия из маргинальных клеток в улитковый проток.
Другой член семейства KCNQ калиевых каналов, KCNQ4, скорее всего, участвует и базолатеральной секреции К+ волосковыми клетками. Мутация у человека гена KCNQ4 приводит к несиндромной глухоте. Другие известные генетические нарушения включают транспортирующие ионы белки с локализацией на базолатеральной мембране краевых клеток. Вероятно, наиболее известные гены, вовлеченные в калиевый гомеостаз улитки — гены, кодирующие белки-коннексины. Коннексины формируют субъединицы щелевых контактов каналов, которые лежат в основе циркуляции К+, описанной для поддерживающих клеток кортиева органа, спиральной связки и сосудистой полоски. Мутация генов, кодирующих коннексины человека 26, 30, 31 и 43, отвечает за большинство несиндромных наследственных потерь слуха.
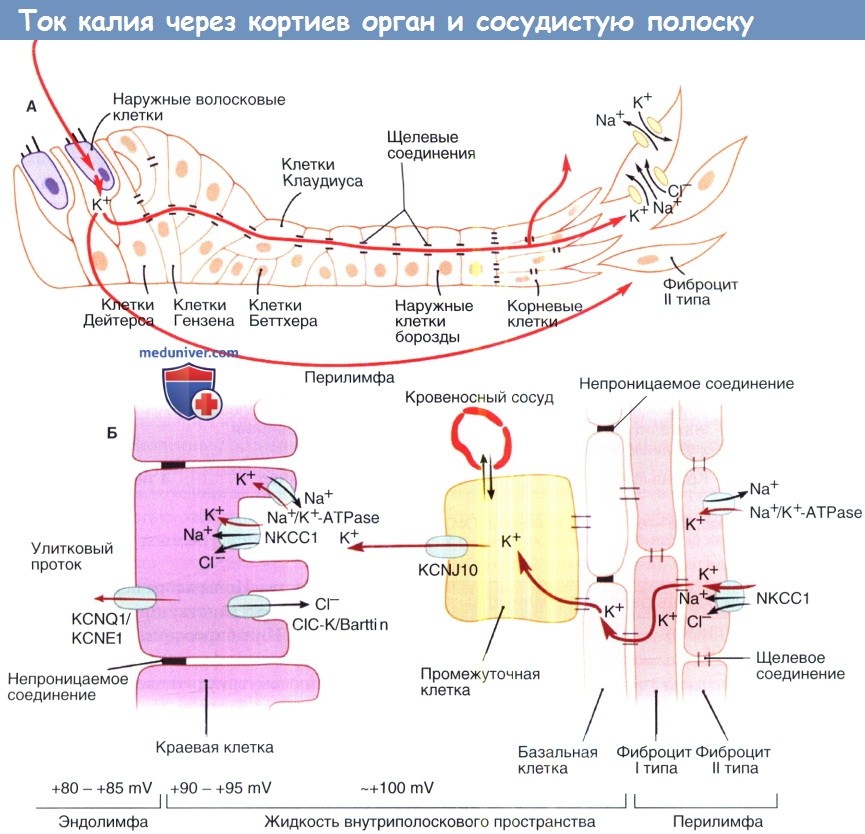
Сосудистая полоска и циркуляция ионов K+.
Схема тока ионов К+ от улиткового протока через волосковые клетки в перелимфатическое пространство,
а также через межклеточные пространства к спиральной связке.
К+ от спиральной связки транспортируется посредством сосудистой полоски в улитковый проток.
Не показаны другие пути тока К+ из улиткового протока, а именно через наружные клетки борозды и мембрану Рейсснера.
Прямоугольной скобкой выделено пространство, более детально показанное на рисунке ниже.
Ток К+ через кортиев орган и сосудистую полоску.
А. К+ входит в волосковые клетки посредством каналов механической передачи.
В базально-латеральных отделах калий выводится в перелимфатическое пространство посредством K+-каналов, таких как канал KCNQ4.
К+ может перемещаться по направлению к спиральной связке через перелимфатическое пространство и внутриклеточно, через сеть межклеточных пространств.
Фиброциты II типа спиральной связки выводят К+ и обеспечивают его передвижение к сосудистой полоске через сеть пространств в соединительной ткани.
Б. К+ входит в базальные и промежуточные клетки через пространства между фиброцитами I и II типов.
К+-канал KCNJ10 является важным звеном в высвобождении калия в пространство внутри сосудистой полоски.
Ген, кодирующий KCNJ10, необходим для правильной генерации внутриулиткового потенциала.
К+ эффективно выводится из пространства внутри сосудистой полоски краевыми клетками, которые активно выводят К+ посредством NKCC1 (Na+/К+/2Cl) ко-транспортеров и Na+/К+-АТФаз.
В итоге, маргинальные клетки секретируют К+ в улитковый проток посредством KCNQ1/KCNE1 К+-каналов поддерживая высокую концентрацию К+ в эндолимфе, необходимую для механоэлектрической передачи.
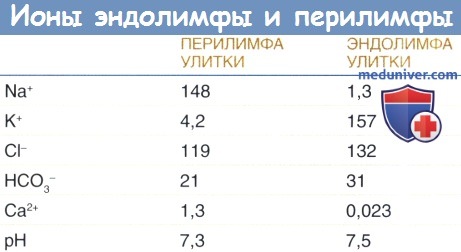
б) Гомеостаз жидкостей улитки. Перелимфа, эндолимфа и жидкость, располагающаяся внутри сосудистой полоски (интрастриарная жидкость), являются тремя разными жидкостями улитки, которые можно рассматривать как систему метаболической поддержки. Для генерации и поддержания внутриулиткового потенциала необходим сбалансированный ионный состав этих трех жидкостей. Перелимфа и интрастриарная жидкость характеризуются высоким содержанием Na+ и низким содержанием К+, как и другие внеклеточные жидкости.
Эндолимфа не только имеет высокую концентрацию К+ и низкую концентрацию Na+, но также имеет необычайно низкую концентрацию ионов Са2+ по сравнению с другими внеклеточными жидкостями. Гомеостаз ионов Са2+ в улитке контролируется ионными каналами и переносчиками, находящихся в плазматических мембранах клеток, как было описано выше для калия.
В сосудистой полоске поступление ионов Na+ сопряжено с выведением ионов К+ из интрастриарного пространства в маргинальные клетки. Переносчики NKCC1 используют высокие градиенты натрия для переноса ионов Na+, К+ и 2Cl- в маргинальные клетки. Na+/K+-АТФ-аза контролирует работу этого градиента посредством нагнетания Na+ в интрастриарное пространство в обмен на К+.
Наконец, К+ покидает маргинальные клетки и выводится в эндолимфатическое пространство под воздействием высокого положительного потенциала покоя маргинальных клеток. Этот сложный процесс поддерживает высокую концентрацию ионов Na+ и низкую концентрацию К+ в интрастриарной жидкости, что облегчает пополнение калием интрастриарного пространства. Ионы хлора переносятся обратно к интрастиарному пространству при помощи С1С-К/барттиновых каналов. Ингибирование NKCC1 и Na+/К+-АТФ-азы петлевым диуретиком фуросемидом и уабаином приводит к уменьшению внутриулиткового потенциала.
Мутация гена, кодирующего барттин, или мутация обеих субъединиц базолатеральных Cl- каналов ClC-Ка и ClC-Kb приводит к синдрому Барттера 4 типа, который характеризуется глухотой и потерей почкой солей. Натрий реабсорбируется из эндолимфы наружной бороздой и клетками Рейснеровой мембраны, которые играют роль в сохранении низкой концентрации Na+ в улитковом протоке.
Регулирование концентрации ионов Са2+ в эндолимфе крайне важно. Изучение волосковых клеток показало, что при низких концентрациях Са2+ происходит разрыв концевых связей, а при высоких концентрациях Са2+ блокируются каналы механоэлектрической передачи. Кроме того, ионы Са2+ частично обеспечивают работу потоковой передачи и играют важную роль в адаптации и, возможно, в улитковой амплификации. Са2+-проницаемые каналы, кальциевая АТФ-аза, так же как и Na+/Ca2+-обмен обнаруживаются в волосковых клетках многих типов и могут участвовать в регуляции притока и оттока Са2+ из эндолимфы, однако точные механизмы этого еще неизвестны.
Регуляция объема жидкостей улитки также важна для функционирования улитки, описано множество различных механизмов, поддерживающих эту регуляцию. Первоначально, в качестве основных принципов были предположены продольные и радиальные потоки. Продольный ток эндолимфы описывался как ее секреция на протяжении перепончатого лабиринта с реабсорбцией в эндолимфатическом протоке и мешке, в то время как радиальный ток формировался локальной секрецией и реабсорбцией, в частности, посредством сосудистой полоски.
За патологические состояния, такие как увеличение или уменьшение объема эндолимфы, может быть ответственен продольный ток. Было обнаружено, что экспериментальное увеличение эндолимфы вызывало продольный ток эндолимфы по направлению к основанию улитки в эндолимфатический мешок, снижая как объем жидкости, так и концентрацию электролитов в улитковом протоке. С другой стороны, экспериментальное уменьшение объема эндолимфы приводило к току, направленному к верхушке, увеличению объема жидкости и концентрации электролитов. Теория радиального тока никогда не была подтверждена экспериментально.
В настоящее время преобладает мнение, что в физиологических условиях не существует значимого тока эндолимфы. Эксперименты на животных показали, что меченые ионы, помещенные в эндолимфу без нарушения ее объема, двигались исключительно посредством диффузии. Таким образом, ионы эндолимфы циркулировали без формирования объемного тока. Схожие регуляторные механизмы предложены для объяснения гомеостаза перелимфы. Медленный объемный ток внутри улитки играет роль при внутриулитковом введении препаратов, где в физиологических условиях диффузия в отдельных наполненных жидкостью отсеках, по всей видимости, равномерно распределяет концентрации препаратов от основания до верхушки.
На клеточном уровне трансмембранное движение воды во многом зависит от напоминающих поры водопроницаемых каналов, таких как аквапорины. Некоторое количество аквапоринов было обнаружено во внутреннем ухе, но лишь немногие из них локализовались в выстилающем эдролимфу эпителии. Снижение аквапорина-4 у мыши приводило к нарушениям слуха. Мыши с другими модифицированными генами аквапорина либо имели неизвестный на сегодняшний день фенотип, либо погибали в эмбриональном периоде, как и мыши с модифицированным геном аквапорина-2. Тем не менее, аквапорин-2 интересен тем, что он обнаруживается в эпителии, выстилающем эндолимфатический мешок, и регулируется гормоном вазопрессином.
Модели животных показали, что патологически повышенный уровень вазопрессина приводил к выраженному отеку, морфологически характерному для болезни Меньера. Помимо потенциальной роли в патогенезе болезни Меньера данный факт дает основания говорить о гормональном влиянии на регуляцию объема жидкостей внутреннего уха. В частности, вазопрессин, как было показано, не только влияет на мембранную экспрессию аквапорина-2, но также увеличивает активность эпителиальных натриевых каналов и NKCC1 переносчиков, обнаруживаемых в маргинальных клетках сосудистой полоски и фиброцитах II типа спиральной связки. Модуляция этих каналов ведет к увеличению секреции калия в эндолимфу, а впоследствии к изменению осмотического объема, приводя к типичному гидропсу. Сходные данные были получены для другого гормона, альдостерона, который увеличивает активность как эпителиальных натриевых каналов, и Na+/К+-АТФ-азы. Однако, другие гормоны имеют противоположный вазопрессину эффект.
Глюкокортикоиды, в частности, как было показано, подавляют симптомы, возникающие при болезни Меньера, возможно, из-за снижения выработки вазопрессина и модуляции мембранной экспрессии определенных аквапоринов.
Гомеостаз жидкостей улитки, ионный транспорт и внутриулитковый потенциал необходимы для правильного функционирования улитки. Метаболическая блокада или специфическое угнетение ионного транспорта, в частности уабаином или фуросемидом, быстро поражает эту микросреду и нарушает слуховую функцию, снижая заряд улитки, то есть внутриулитковый потенциал. Есть предположение, что старение влияет на внутриулитковый потенциал и возникающее вслед за этим снижение метаболизма сосудистой полоски может играть роль в возрастной потере слуха.