Улитка является органом слуха млекопитающих, заключена в височную кость и представляет собой костную структуру по форме напоминающую раковину улитки («cochlos» — греч. «улитка»). У людей улитка закручена в 22/3 оборота вокруг центральной оси, модиолуса. В костной части улитки (также известной как костный лабиринт), имеются каналы или лестницы, образованные перепончатым лабиринтом: центральная, срединная лестница, также известная как улитковый проток, отделена от вестибулярной лестницы преддверной мембраной (мембраной Рейсснера) и от барабанной лестницы базальная мембраной.
Вестибулярная лестница соединяется со средним ухом через овальное окно, которое прилегает к подножной пластинке стремени. Круглое окно, соединяющее барабанную лестницу и среднее ухо, прикрыто мембраной круглого окна. Вестибулярная и барабанная лестница соединяются на вершине улитке (в области геликотремы); улитковый проток заканчивается слепо. Вестибулярная и барабанная лестницы наполнены перилимфой, внеклеточной жидкостью с высоким содержанием Na+ и низким содержанием К+, в то время как улитковый проток наполнен эндолимфой, отличающейся высоким содержанием К+ и низким содержанием Na+. Эндолимфа улитки имеет положительный электрический потенциал, который приблизительно равен +85 мВ.
Разница в концентрации ионов между перелимфой и эндолимфой и разность потенциалов обеспечивает энергию, необходимую для работы улитки.
С физиологической точки зрения в улитке можно выделить три функциональных отдела:
(1) кортиев орган, представляющий «датчик» улитки, преобразующий механические колебания в электрические сигналы (механоэлектрическая передача);
(2) сосудистую полоску, улитковую «батарею», генерирующую энергию (эндокохлеарный потенциал), для механоэлектрической передачи и влияющую на гомеостаз жидкостей улитки;
(3) спиральный ганглий, включающий аксоны нейронов («электрический провод»), передающий электрические сигналы от улитки к центральной нервной системе.
Все три отдела необходимы для нормального функционирования улитки и будут детально описаны в отдельной статье на сайте (список представлен в конце статьи, либо просим пользоваться формой поиска на главной странице сайта).
Кортиев орган является рецептором органа слуха млекопитающих. Так он был назван после того, как в XIX веке, итальянский гистолог Альфонсо Корти, впервые обнаружил и описал этот морфологически сложный орган.
Кортиев орган состоит из двух типов чувствительных рецепторов, внутренних и наружных волосковых клеток. Около 3500 бокаловидных внутренних волосковых клеток выстроились в один ряд по всей длине улиткового протока. Латеральнее внутренних клеток обнаруживаются три ряда наружных волосковых клеток, отличающихся специфической цилиндрической формой.
Оба типа волосковых клеток представляют собой пучки из высокоорганизованных, содержащих актин стереоцилий с увеличением высоты к латеральному ряду, таким образом, самые короткие находятся в середине. Волосковые пучки волосковых клеток внутреннего уха образуют изогнутую линию, состоящую из 2-3 рядов стереоцилий. Пучки наружных волосковых клеток имеют вид буквы V. Пучки волосковых клеток являются их механочувствительными органеллами.
Каждая волосковая клетка располагается на верхушке фаланговой клетки, которая у наружных волосковых клеток называется клеткой Дейтерса. Внутренние и наружные столбовые (поддерживающие) клетки условно разделяют пространство между внутренними и наружными волосковыми клетками и образуют туннель кортиева органа. Также имеются и другие поддерживающие клетки кортиева органа. С медиальной стороны расположены внутренние пограничные клетки, латеральнее — клетки Гензена (кнаружи от пограничных клеток), клетки Клаудиуса, и клетки Беттхера. По всей своей длине кортиев орган покрыт покровной мембраной. Эта бесклеточная структура, внутренней частью укрепленная на преддверной губе спиральной пластинки улитки и соединенная с пучками наружных волосковых клеток.
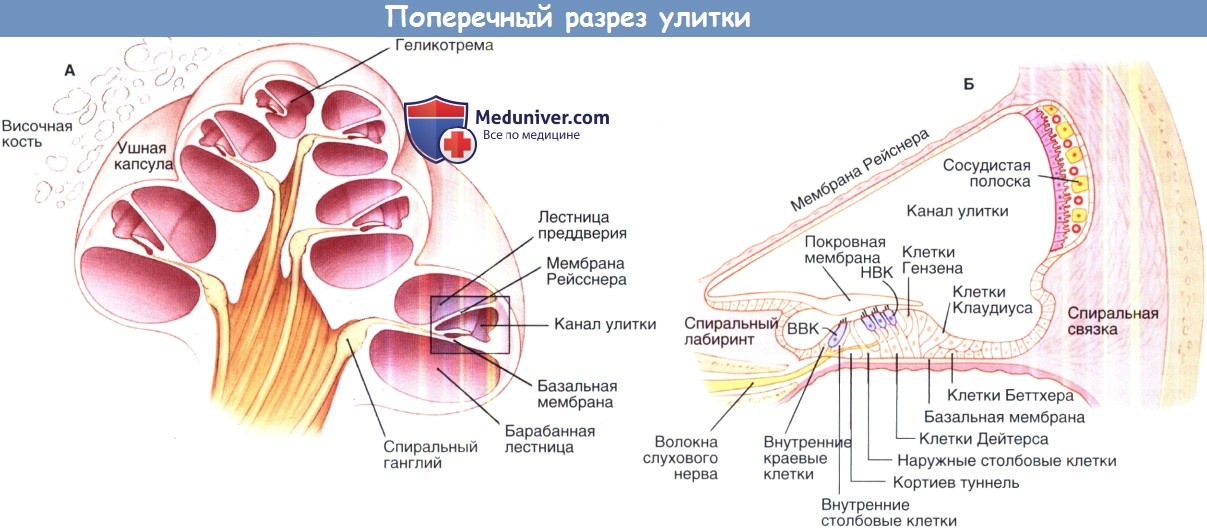
Поперечный разрез улитки. А, схематичное изображение улитки в разрезе.
Выделенная область, показанная на рисунке Б, изображает улитковый проток и окружающие структуры, такие как кортиев орган и сосудистую полоску в разрезе.
ВВК — внутренние волосковые клетки; НВК — наружные волосковые клетки.
а) Базиллярная мембрана и тонотопия кортиева органа. Когда звук воздействует на барабанную перепонку, вибрация передается на внутреннее ухо через три слуховые косточки. Движение стремени вызывает смещение жидкости улитки в вестибулярной лестнице. Несжимаемость перелимфы создает градиент давления между вестибулярной и барабанной лестницами, приводя в движение базальную мембрану и кортиев орган. Это смещение может быть представлено в виде бегущей волны, которая движется от основания к верхушке вдоль базальной мембраны. Для стимулов чистых тонов бегущая волна достигает максимума в определенном месте базальной мембраны и затем спадает.
Точное месторасположение этого максимума зависит от частоты стимула, что является основополагающим принципом тонотопической организации в улитке. Специфическое влияние частоты на определенные участки базальной мембраны регулируется свойствами всех пассивных компонентов, таких как внеклеточные, клеточные и молекулярные структуры в этом месте, а также свойствами активных систем, таких как кохлеарный усилитель (описан далее). Основание улитки у людей настроено на частоты до 20 кГц, а в области верхушки на частоты до 20 Гц. Тонотопический градиент анатомически проявляется не только в изменении ширины базальной мембраны, но и в изменениях высоты волосковых клеток и длинны клеточных структур, таких как стереоцилии и собственно пучки волосковых клеток.
Внутренние волосковые клетки улитки служат основой слуха, являясь сенсорными клетками, преобразующими механические стимулы в электрические сигналы и посредством синаптической активности передающими эти сигналы в мозг. В основе этого процесса лежит механоэлектрическая передача, которая возникает на самом конце или около кончика стереоцилии. Упомянутый аппарат механоэлектрической передачи присутствует во всех волосковых клетках и состоит из одного или более механически закрытых катионных каналов, тесно связанных эластичных структур и концевой связью стереоцилии со следующей, более высокой стереоцилией.
Механические отклонения стереоцилий волосковых клеток в сторону более высоких рядов приводит к смещению соседних стереоцилий. Постепенное увеличение механического напряжения в передаточном аппарате ведет к конформационным изменениям белка-передатчика в канале и к увеличению пропускной способности канала, которая в покое у млекопитающих составляет 40-50%. Несмотря на некоторое количество белков-кандидатов, ни один из предполагаемых компонентов передаточного аппарата волосковых клеток не был однозначно функционально связан с биофизическим процессом механической передачи. Наиболее вероятным кандидатом является кадхерин-23 и протокадхерин-15, которые были предложены в качестве компонентов концевой связи, а также миозин-1с, необходимый для адаптационного процесса, контролирующего заданное значение механочувствительности.
Мутация гена, кодирующего либо кадхерин-23, либо протокадхерин-15 у человека, приводит к синдрому Ушера (врожденной глухоте с прогрессирующей потерей зрения в результате пигментного ретинита).
При механической стимуляции в направлении самого высокого ряда стереоцилий ионы К+ и Са2+ входят в волосковые клетки через открытые каналы механоэлектрической передачи, находящиеся около верхушек стереоцилий. Это отклонение ведет к деполяризации клетки. При отклонении стереоцилии, канал передачи закрывается, тем самым гиперполяризируя клетку. После длительного сгибания пучков волосковых клеток происходит адаптация изначально большой пропускной способности канала, проявляясь уменьшением тока в связи с закрытием передаточных каналов. Было сделано предположение, что за адаптацию отвечают два различных процесса: быстрое закрытие каналов передачи и скольжение миозина, связанно с передаточным аппаратом. Быстрое повторное включение каналов или «быстрая адаптация» предположительно связана с Са2+ во внутриклеточном пространстве около ворот канала. Точный механизм этого процесса пока неясен.
Второй процесс, «медленная адаптация», происходит примерно в 10 раз медленнее, чем быстрое открытие канала, и возникает при соскальзывании концевого соединения стреоцилии вниз. Во время стимула адаптация ведет к переустановке точки покоя, тем самым позволяя передаточному аппарату непрерывно функционировать в области наивысшей чувствительности. Предполагается, что поступление Са2+ через открытые каналы ведет к проскальзыванию входящего в адаптационный аппарат миозина, который смещается по направлению к вершине стереоцилии вдоль актиновых нитей. Проскальзывание миозина уменьшает напряженность на концевом соединении и снижает пропускную способность передаточного канала, которая в свою очередь, выключает местный приток Са2+.
При низких концентрациях Са2+ миозин адаптационного аппарата будет эффективно двигаться вверх, тем самым восстанавливая напряженность концевой связи до точки, когда пропускная способность передаточного канала будет близка к пропускной способности в покое. Миозин 1с выдвигается в качестве важного компонента адаптационного механизма, что не исключает участия других типов миозина в этом процессе.
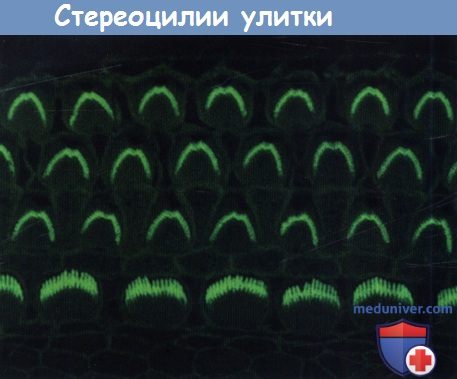
Стереоцилии улитки. Показан апикальный участок кортиева органа, длиной 150 мкм.
Пучки стереоцилий окрашены фаллоидином, меченым флюоресцеином, который связывается с филаментами актина.
Изогнутые пучки внутренних волосковых клеток находятся внизу, наверху видны три ряда V-образных пучков наружных волосковых клеток.
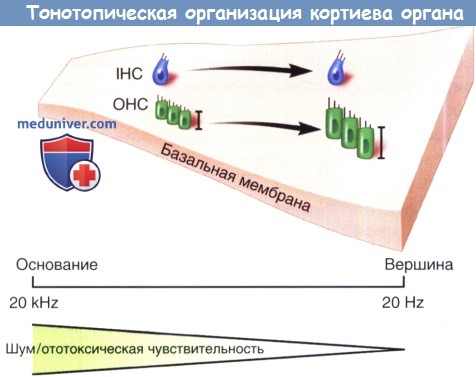
Тонотопическая организация кортиева органа.
Схематическое изображение анатомических изменений на протяжении улитки от основания к верхушке,
включающее увеличение ширины базальной мембраны и размера наружных волосковых клеток.
Эти изменения обеспечивают перестройку частоты кортиева органа.
Подобным образом чувствительность к ототоксическому повреждению, такому как шум или действие аминогликозидов, выше в основании улитки и уменьшается по направлению к верхушке.
IHC — внутренние волосковые клетки; ОНС — наружные волосковые клетки.
б) Наружные волосковые клетки и амплификация. Наружные волосковые клетки играют ключевую роль в амплификации движений базальной мембраны. Амплификация (усиление) необходима для улавливания звуков с низким уровнем звукового давления. Важная роль наружных волосковых клеток была продемонстрирована в случаях, когда канамицин (ототоксичный антибиотик) использовался для селективного повреждения наружных волосковых клеток, в то время как внутренние волосковые клетки оставались интактными. Повреждение наружных волосковых клеток приводит к исключению низкого порога чувствительности слухового нерва и его точной настройки, но также влияет и на характеристики высокого порога. Это наблюдение привело к гипотезе, что наружные волосковые клетки являются несущими главную ответственность за амплификацию и точную настройку слуховой системы.
Одним из механизмов усиления является соматическая электроподвижность. Длина изолированных наружных волосковых клеток в ответ на электростимуляцию изменялась на 3-5%. Во время деполяризации наружные волосковые клетки сокращались, в то время как при гиперполяризации возникало их удлинение. В результате наружные волосковые клетки оказывают механическое воздействие, которое приводит базальную мембрану в движение при стимулах до нескольких кГц.
Усиление стимуляции базальной мембраны, в частности, движение стереоцилий в направлении возбуждения и деполяризация наружных волосковых клеток, которые в свою очередь сокращаются, подтягивает базальную мембрану кверху. Таким образом электроподвижность наружных волосковых клеток усиливает подвижность базальной мембраны, вызванную бегущей волной.
Полагают, что престин является белком, ответственным за соматическую электроподвижность в наружных волосковых клетках. Это подтверждается рядом доказательств. Во-первых, при трансфекции престином клетки демонстрируют электроподвижность с амплитудой (размером) до 0,2 мкм, что говорит о необходимости перстина для движения. Во-вторых, престин обнаруживается в нужном месте, а именно в латеральной мембране волосковых клеток. И, наконец, прецизионное удаление или модификация престина у мышиной модели влияло на чувствительность улитки и указывало на необходимость престина для электроподвижности наружных волосковых клеток. Престин относится к суперсемейству передачи анионов SCL26, которые могут быть посредниками электронейтрального обмена хлорида и карбоната через плазматическую мембрану.
Точный механизм, благодаря которому работает этот насос, по-прежнему неизвестен, однако можно предположить, что белковый насос, работающий на принципах изменения напряжения, способен работать гораздо быстрее, чем классическая АТФ-зависимая помпа. В настоящее время, согласно рабочей гипотезе, внутриклеточные анионы действуют как датчики напряжения, связанные с престином и запускающие конформационные изменения. Гиперполяризация ведет к связыванию анионов с престином, что вызывает увеличение поверхности престина, и приводит к увеличению клетки. С другой стороны, деполяризация ведет к отсоединению анионов, уменьшению поверхности престина и к сокращению клетки. В покое анионы обычно отделены от престина, поэтому белок может трансформироваться.
Другой вероятный источник амплификации связан с активным движением пучка волосковых клеток, вызванного воздействием механической передачи и адаптации. В улитке не относящихся к млекопитающим позвоночных пучки волосковых клеток способны генерировать устойчивые колебательные движения, подобная сеть усиления присутствует в наружных волосковых клетках млекопитающих. Активные движения стереоцилий являются важным механизмом амплификации у немлекопитающих позвоночных, и вполне вероятно, что подобный процесс также используется для усиления или настройки кортиева органа у млекопитающих в тесной связи с электроподвижностью наружных волосковых клеток, управляемой престином.
в) Покровная мембрана кортиева органа. Покровная мембрана является внеклеточной структурой, которая лежит на внутренних и наружных волосковых клетках. Однако, только наиболее высокие стереоцилии волосковых клеток непосредственно вплетены в ее нижнюю часть. Покровная мембрана прикрепляется к внутреннему краю лимба и подвижно связана с поддерживающими клетками, такими как клетка Гензена, посредством микроскопических тяжей, названных трабекулами. Важность покровной мембраны подтверждается тем, что мутация гена, кодирующего покровную мембрану (альфа- и бета-текторин), является причиной выраженной потери слуха как у человека, так и у животных.
Исходя из анатомических наблюдений, вначале полагали, что покровная мембрана действует как простой рычаг, который двигает пучки волосковых клеток, перемещаясь вверх и вниз. Другие модели улитки рассматривали мембрану как простой механический груз или резонансную систему, обладающую массой и упругостью. Недавние исследования показали, что покровная мембрана больше похожа на резонансный гель и участвует в увеличении частотной чувствительности улитки. Вполне вероятно, что все предложенные функции покровной мембраны являются актуальными и так как эта структура, как и большинство структур кортиева органа, изменяет свой размер и форму от основания к вершине, покровная мембрана может также способствовать общей тонотопической организации улитки.
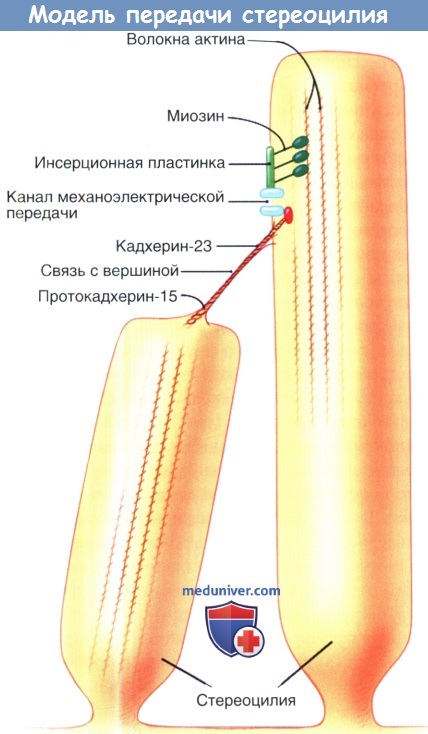
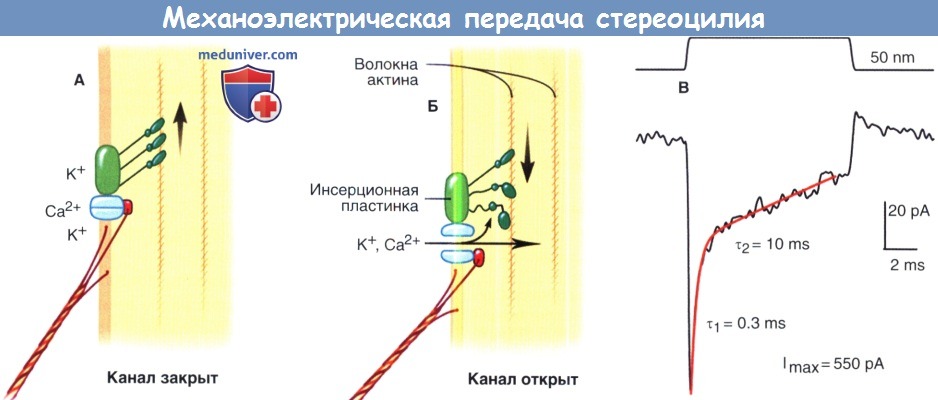
Модель аппарата механоэлектрической передачи стереоцилии.
Изображены известные и предполагаемые компоненты аппарата механоэлектрической передачи.
Механоэлектрическая передача.
А. В покое приблизительно 90% каналов передачи закрыты. Молекулярные насосы, основанные на миозине,
поднимаются к вершине стереоцилии и регулируют натяжение концевой связи и связанные с ней структуры для обеспечения более точной чувствительности передаточного аппарата.
Б. увеличение механического натяжения в области концевой связи и связанных с ней структур ведет к открытию каналов передачи и входу катионов, деполяризующих клетку.
Местное увеличение концентрации ионов Са2+ активирует миозиновые насосы и приводит к скольжению передаточного аппарата, тем самым уменьшается механическое натяжение и открываются каналы передачи.
В. деполяризация наружных волосковых клеток крысы в ответ на умеренное механическое отклонение в 50 нм.
Показан быстрый рост рецепторного потенциала, который позволяет достигнуть максимума в 550 пА в отдельно взятой клетке при максимальной стимуляции.
Представленная схема отображает время быстрой (τ1) и медленной (τ2) адаптации.