Сроки и стадии нормального развития центральной нервной системы: эмбриогенез ЦНС
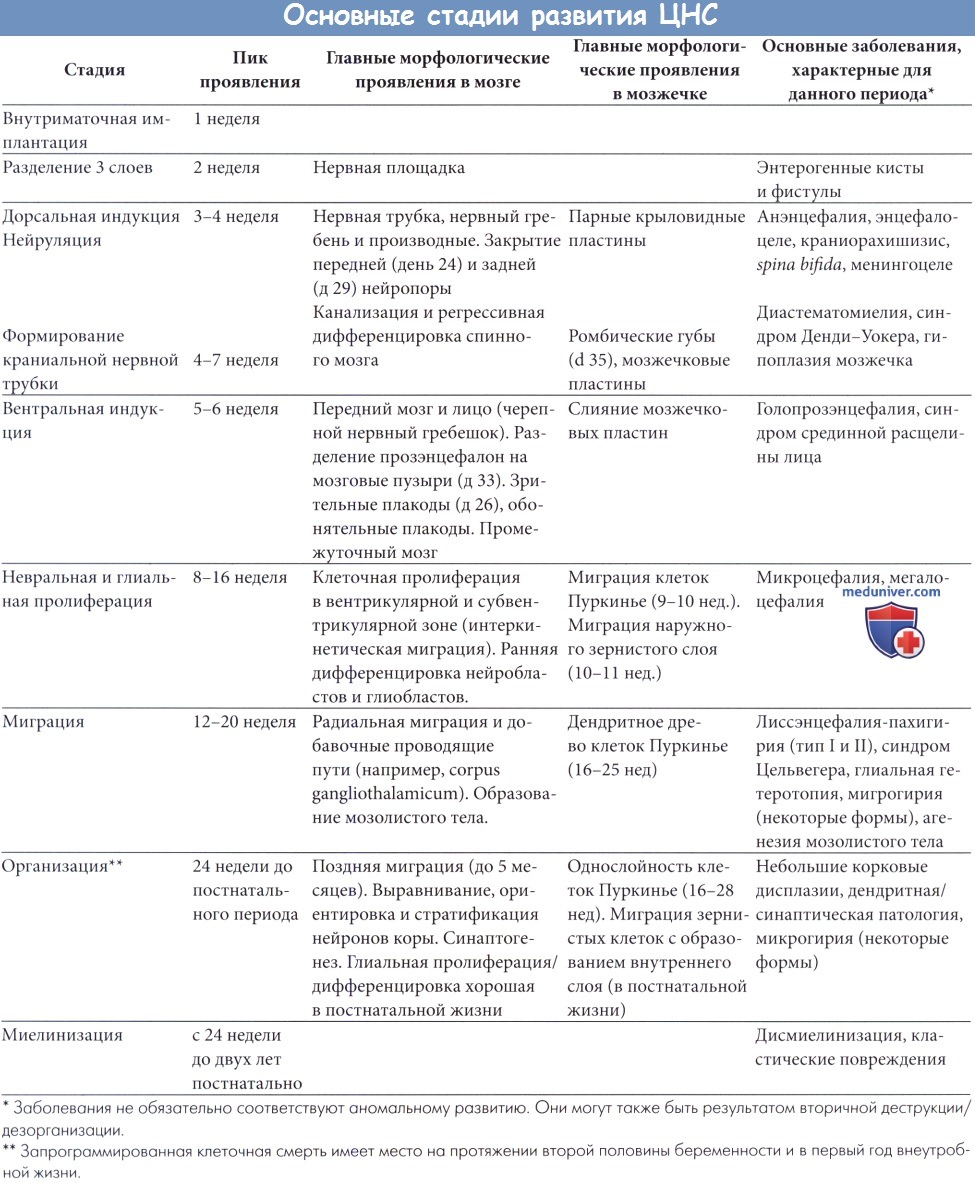
Рассмотрим вкратце эмбриогенез ЦНС. Более полная информация содержится в классических работах (Sarnat, 1987, Barkovich et al., 1992, McConnell, 1992). В таблице ниже представлен схематический обзор основных стадий эмбриогенеза ЦНС. Регуляция развития ЦНС, фантастически сложного процесса, контролируется многими факторами, особенно генетическими. Главную роль играет большой набор белков, кодируемых или регулируемых генами (гомеобокс-гены и факторы транскрипции), которые определяют градиент дифференцировки (переднее-задний и дорсовентральный) и ограничивают определенность основных организационных единиц. Эти механизмы до сих пор в значительной степени неясны, и их изучение выходит за рамки данной книги.
На протяжении второй недели эмбриогенеза формируются три слоя из эктодермы, мезодермы и энтодермы. Через две недели срединная часть эктодермы, под влиянием подлежащей мезодермы, становится нервной пластинкой, которая в дальнейшем развивается в нервный желобок, а затем в нервную трубку. На четвертой неделе нервная трубка закрывается. Процесс замыкания начинается в средней части трубки и направляется к конечностям. Молекулы адгезии нервных клеток играют центральную роль в процессе закрытия (Sarnat и Flores-Sarnat, 2002, 2004).
Замыкание нервной трубки определяется дорсальной индукцией со стороны мезодермы. До закрытия трубки на девятый день на переднем конце эмбриона становятся различимыми зачатки будущего ромбэнцефалона, мезэнцефалона, прозэнцефалона и ушные плакоды. Одновременно, группы клеток вдоль боковых краев нервной трубки отделяются и формируют парные невральные гребни, которые дают начало основным структурам периферической нервной системы, оболочкам и меланоцитам.
Задняя часть нервной пластины и хорда имеет различный исход: из нее формируется масса клеток, в которых в дальнейшем произойдет канализация и процесс регрессивной дифференцировки с образованием нижнего уровня спинного мозга.
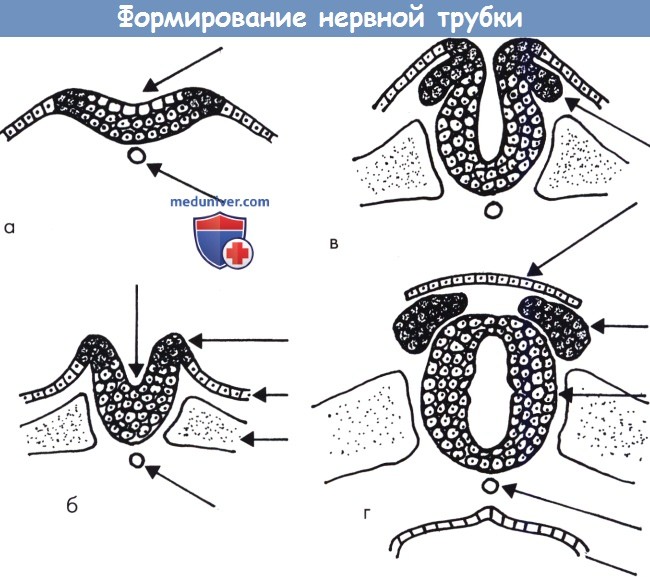
Формирование нервной трубки (схема):
(а) Нервная пластинка формируется из утолщенной эктодермы под влиянием хорды.
(б) Возникает нервный желобок. Видны зачатки нервного гребня.
(в) Нервный гребень хорошо сформирован и его клетки мигрируют в латеральном направлении к своим мишеням.
(г) Закрытие нервной трубки начинается в центральной области; концы (передние и задние нейропоры) закрываются позже.
Закрытая трубка покрыта эктодермой и мезенхимальной тканью.
К 32-33 дню в переднем мозговой пузыре формируются телэнцефалические пузыри и дифференцируется промежуточный мозг, так что на восьмой неделе представлены четыре определяемые клеточные массы в области базальных ядер. Процесс образования пузырей, по всей видимости, происходит под влиянием вентральной индукции хорды, но точный механизм индукции до конца непонятен.
Приблизительно с 30 дня основные индуктивные процессы заканчиваются и начинается клеточная дифференцировка. Размножение примитивных клеток, которые затем становятся как нейронами, так и клетками глии, происходит, главным образом, около полости желудочков и, в меньшей степени, в субвентрикулярной зоне. В глубоких клеточных слоях встречаются митозы (Caviness et al., 2003).
Деление клеток является асимметричным: одна дочерняя клетка мигрирует к внешней стороне во время телофазы, затем возвращается в глубокую область, чтобы начать следующий цикл; так называемая интракинетическая миграция остается в желудочковой зоне. Другая клетка покидает желудочковую зону, чтобы начать свое путешествие к кортикальной пластинке.
Пролиферация нейробластов достигает максимума к 15 неделе гестации, снижаясь затем до остановки на 20 неделе. Пролиферация тесно связана с плохо понимаемым процессом запрограммированной клеточной смерти через апоптоз. Активность процессов апоптоза повышается в то же время, когда пролиферация уменьшается. Фактическая пропорция умирающих клеток у человека точно неизвестна. Вероятнее всего, этот процесс варьирует в зависимости от локализации и может в некоторых областях затрагивать более 30-50% сформировавшихся клеток.
Кортикогенез частично изучен за последние два десятилетия. В позднем эмбриональном периоде (45-50 дни гестации) примитивные кортикопетальные волокна проникают через диэнцефальную борозду и распространяются под телэнцефальной мягкой мозговой оболочкой; нейроны, по всей видимости, в пределах этих волокон формируют зародышевый плексиформный слой (ИПС) (Bentivoglio et al., 2003) или препластинку. ИПС предшествует миграции нейронов, формирующей кортикальную пластинку, и служит опорой мигрирующим клеткам.
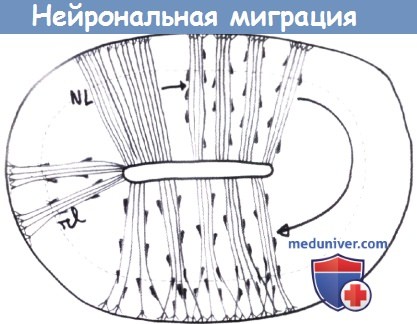
Схематическое изображение нейрональной миграции вдоль радиальных глиальных проводников. Поперечное сечение нервной трубки (с разрешения Р. Evrard, Cliniques Saint-Luc).
NL: Эмбриональная стадия — равномерно распределенные глиальные проводники, первая стрелка: миграция нейронов направлена к глубоким кортикальным слоям вдоль глиальных проводников,
в это время сгруппированных в пучки, изогнутая стрелка: поздние стадии нейрональной миграции (слои II и III).
Прогрессирующая дефасцикуляция глиальных проводников.
rl: Картина у «шатающихся» мутантных мышей, у которых не происходит дефасцикуляции.
Мигрирующие нейроны делят корковую зону на поверхностную часть, которая будет составлять молекулярный слой или слой I, в основном содержащий клетки Кахаля-Ретциуса и их отростки (Sarnat и Flores-Sarnat, 2002, Rakic и Zecevic, 2003) и глубокий слой, называемый субпластинкой (или слоем VII), который исчезает до окончания внутриутробной жизни. Клетки КахаляРетциуса играют важную роль в окончательной судьбе пирамидальных клеток, вероятно, с помощью секретируемого рилина (Crino, 2001, Assadi et al, 2003). Большинство из них исчезает за счет апоптоза в возрасте до одного года. Субпластинки нейрона действуют как «переключатель» для кортикопетальных талямических аксонов до того, как клетки четвертого слоя начнут функционировать. Поздние мигрирующие клетки, которые сформируют слои со II по VI дефинитивной коры, перемещаются способом «наизнанку», то есть молодые генерированные поздне-мигрирующие клетки образуют более поверхностные слои (слой II) корковой пластины, в то время как большие пирамидные клетки слоев V и VI мигрировали раньше.
Большинство нейронов будущей коры мозга перемещается между 10-й и 18-й неделями гестации, и полный комплекс корковых нейронов по существу завершается к 20-й неделе. Однако клеточная миграция продолжается в более медленном темпе на протяжении беременности, и некоторые клетки, например, зернистые мозжечковые и гиппокампальные, мигрируют постнатально (Sarnat, 1987). Некоторый корковый нейрогенез сохраняется в течение жизни, особенно в средней височной доле и мозжечке (Gage, 2002).
Нейрональная миграция является сложным процессом, который отличается в зависимости от типов нейронов. Радиальная миграция затрагивает большинство (вероятно, 75%) клеток-предшественниц, предназначенных для образования пирамидных нейронов. Они мигрируют вдоль глиальных проводников, которые простираются от желудочковой (пролиферативной) зоны к пиальной поверхности нервной трубки и производных структур (Rakic, 1981, Williams и Caviness, 1984). Глиальные проводники позднее превращаются в астроциты, завершая радиальную миграцию. Несколько клеток используют одни и те же глиальные волокна при перемещении, что может отвечать за модульную организацию коры, единую для млекопитающих (Rockel et al., 1980), модули представляют собой функциональные единицы коры. Функциональные единицы видов млекопитающих отличаются количеством, а не структурой. Не все нейроны следуют точно за особым глиальным проводником.
Другие клетки-предшественницы, которые затем станут интернейронами, отделяются от проводника и мигрируют в перпендикулярном направлении (Walsh, 1995, Caviness et al., 2003), часть может направиться к соседним проводникам. Некоторые клетки зарождаются в ганглионарных возвышениях в базальном мозге и мигрируют тангенциально (Jimenez et al., 2002, Bystron et al., 2005, Kanatani et al., 2005). Они следуют по до сих пор плохо известному маршруту, возможно, вдоль аксональных пучков к корковой пластине. Большинство, вероятно, интернейроны, в основном ГАМК-эргические, тогда как пирамидальные (глутаматэргические) клетки мигрируют радиально. Эти разнообразные маршруты могут объяснить широкое распространение в коре многочисленных постмиграционных нейронов, образованных из одного клона (Walsh и Серко, 1993). Мигрирующие нейробласты направляются к своим окончательным местам рядом сигнальных систем, особенно биохимическими, которые контролируются многочисленными генами, в настоящее время интенсивно изучаемыми (Sarnat и Flores-Sarnat, 2002, Crino, 2004).
Вторая половина гестации характеризуется быстрым увеличением длины и сложности дендритов и аксонов посредством установления синапсов, с созреванием и тонкой организацией коры. В результате быстро увеличивается вес мозга и процесс образования борозд, необходимый для размещения интенсивно увеличивающейся площади коры. Борозды второго и третьего порядка возникают между 7-м и 9-м месяцами беременности, и большинство извилин присутствует в 28 недель. Полная ламинарная структура коры формируется к моменту рождения. Завершающим формирование центральной нервной системы моментом является снижение количества и плотности синапсов в результате развития нейрональных процессов и запрограммированной апоптозом клеточной смерти.
Дендриты, аксоны и синапсы развиваются с огромной скоростью, и многие ранее образованные синусы в итоге исчезают. Развитие дендритов, аксональное ветвление и формирование дендритных корешков продолжается до четвертого года жизни. Глиальное развитие является сложным процессом, разнообразные предшественники имеют неодинаковый исход. Некоторые превращаются в радиальные глиальные волокна, используемые мигрирующими нейронами. Многие астроциты образуются из персистирующей перивентрикулярной пролиферирующей зоны после окончания процессов миграции (Gressens et al., 1992). Миелин начинает образовываться приблизительно в 30 недель, но большая часть процесса образования миелина происходит в постнатальном периоде (Yakovlev и Lecours, 1967, Brody et al., 1987).