Отличительные признаки рака - кратко с точки зрения внутренних болезней
Образование и рост злокачественной опухоли представляют собой многоэтапный процесс, в ходе которого последовательно возникающие генные мутации приводят к образованию раковой клетки. Для запуска процесса канцерогенеза клеткам необходимо обладать характеристиками, которые в совокупности называются отличительными признаками злокачественных новообразований. Отличительных признаков рака ровно 10. Начнем.
1. Нестабильность генома и мутации. Случайные генетические мутации непрерывно происходят во всех клетках организма и очень редко обеспечивают отдельным клеткам избирательное преимущество, позволяющее им непрерывно делиться и доминировать в локальном тканевом окружении.
Многоэтапный канцерогенез обусловлен последовательной клональной экспансией предраковых клеток, причем триггером для каждой такой экспансии является случайная генетическая мутация. В норме механизмы репарации клеточной ДНК настолько эффективны, что почти все спонтанные мутации корректируются, не вызывая фенотипических изменений, поэтому частота мутаций остается очень низкой. В раковых клетках накопление мутаций может быть ускорено вследствие нарушения работы систем наблюдения, которые обычно контролируют целостность генома и вызывают в генетически модифицированных клетках процессы старения или апоптоза.
Именно поэтому эти клетки могут стать более чувствительными к мутагенным воздействиям или в них могут нарушаться механизмы репарации ДНК.
2. Сопротивление клеточной смерти. Существует три основных механизма, которые приводят к гибели клеток в здоровых тканях: апоптоз, аутофагия и некроз.
- Апоптоз. Это запрограммированная гибель клеток. Скорость апоптоза часто существенно замедлена в раковых клетках, особенно в клетках опухолей с высокой степенью злокачественности или резистентных к лечению. Механизм апоптоза имеет регулирующие элементы, которые воспринимают внутренние и внешние проапоптотические сигналы и инициируют каскад протеолиза и разрушения клеток с фрагментацией ядра, конденсацией хромосом и уменьшением размеров клетки с утратой межклеточного контакта, последующей фрагментацией клетки и образованием апоптотических телец, которые фагоцитируются соседними клетками.
Наиболее важным регулятором апоптоза является ген-супрессор злокачественных опухолей ТР53, часто называемый хранителем генома, поскольку он способен индуцировать апоптоз в ответ на значительное повреждение генома. Крупнейшим инициатором апоптоза через ТР53 является повреждение клеток, в частности, вследствие разрушения ДНК на фоне химиотерапии, окислительного повреждения и ультрафиолетового излучения.
- Аутофагия. Это катаболический процесс, в ходе которого клеточные компоненты разрушаются лизосомами внутри самой клетки. Аутофагия — важный физиологический механизм. Обычно он вяло проистекает в клетках, но может быть активирован в ответ на стрессовые факторы окружающей среды (например, на фоне проведения лучевой терапии или цитотоксической химиотерапии). При активации аутофагия выполняет защитную функцию для злокачественных клеток, нивелируя, а не усиливая разрушительные действия этих стрессовых ситуаций. Показано, что клетки злокачественных новообразований, подвергшихся сильному стрессу, с помощью аутофагии уменьшаются до состояния обратимого покоя.
- Некроз. Это преждевременная гибель клеток, характеризующаяся высвобождением клеточного содержимого в местное тканевое микроокружение, в отличие от апоптоза, при котором клетки постепенно разрушаются, а их фрагменты подвергаются фагоцитозу. Некротическая гибель клеток приводит к активации клеток зоны воспаления, стимуляции ангиогенеза и высвобождению стимулирующих факторов, которые увеличивают пролиферацию клеток и инвазию в ткани, тем самым усиливая, а не ингибируя канцерогенез.
3. Поддержание пролиферативной сигнализации. Злокачественные клетки могут поддерживать пролиферацию на более высоком уровне, чем нормальные клетки. Как правило, это происходит из-за факторов роста, которые способны связываться с рецепторами на поверхности клеток, активируя опосредованный тирозинкиназой внутриклеточный каскад передачи сигналов. Это в конечном итоге приводит к изменениям в экспрессии генов и способствует пролиферации и росту клеток.
Устойчивая пролиферативная способность может быть результатом чрезмерного образования лигандов или рецепторов факторов роста и продукции структурно измененных рецепторов, которые могут инициировать сигналы в отсутствие связывания лиганда и без активации компонентов внутриклеточного пути передачи сигналов таким образом, что передача сигналов становится независимой от лиганда.
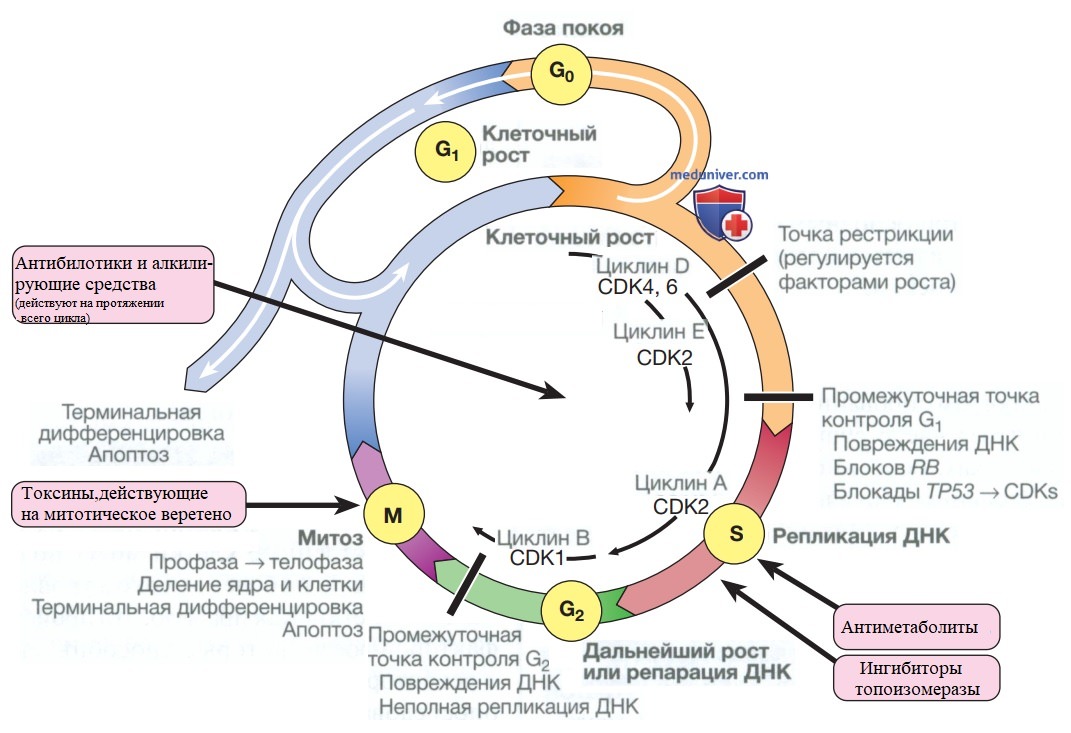
- Клеточный цикл. Клеточный цикл состоит из четырех упорядоченных, строго регулируемых фаз, называемых G1 (пресинтетическая), S (синтетическая, синтез ДНК), G2 (постсинтетическая) и М (митотическая) (рис. 1). В норме клетки, выращенные в культуре, прекратят пролиферацию и перейдут в состояние покоя, называемое G0, как только они сольются или будут лишены плазменных факторов или факторов роста. Пресинтетическая фаза (G1) до начала синтеза ДНК представляет собой промежуточный период, который разделяет митотическую (М) и синтетическую (S) фазы, когда клетки готовятся к репликации ДНК.
Рисунок 1. Клеточный цикл и точки приложения эффектов химиотерапевтических препаратов. CDK — циклинзависимая киназа; RS — ген ретинобластомы
Клетки в фазах G0 и G1 чувствительны к сигналам роста, но как только они прошли точку рестрикции, они без дополнительной стимуляции начинают синтез ДНК (фаза S). В ответ на различные сигналы ингибирования роста клетки могут останавливаться в различных точках фазы G1. Митогенные сигналы способствуют переходу через фазу G1 в фазу S при помощи фосфорилирования продукта гена ретинобластомы (pRB). После синтеза ДНК наступает вторая промежуточная фаза (G2) перед митозом (М), позволяющая клеткам исправить ошибки, которые произошли во время репликации ДНК, и таким образом предотвратить передачу этих ошибок в дочерние клетки.
Хотя продолжительность отдельных фаз может различаться в зависимости от типа клеток и тканей, большинство взрослых клеток в любой отдельный момент времени находятся в состоянии G0.
- Регуляция клеточного цикла. Клеточный цикл упорядочен за счет действия ряда молекулярных механизмов, к наиболее важным из которых относят циклины и циклинзависимые киназы (cyclin-dependent kinases). Циклины связываются с циклинзависимыми киназами и регулируются как активирующим, так и инактивирующим фосфорилированием, с двумя основными контрольными точками при переходе G1/S и G2/M. Гены, ингибирующие прогрессирование, играют важную роль в профилактике опухолей и называются генами-супрессорами опухолей (например, гены ТР53, ТР21, ТР16).
Продукты этих генов дезактивируют комплексы «циклин — циклинзависимые киназы » и таким образом способны останавливать клеточный цикл. Сложность контроля клеточного цикла повышает чувствительность к дисрегуляции генной функции, что может привести к появлению злокачественного фенотипа.
- Стимуляция клеточного цикла. Многие раковые клетки синтезируют факторы роста, которые управляют своей собственной пролиферацией благодаря положительной обратной связи, что носит название аутокринной стимуляции. К ним относят трансформирующий фактор роста α и тромбоцитарный фактор роста (platelet-derived growth factor). Другие раковые клетки избыточно экспрессируют рецепторы к факторам роста вследствие амплификации генов или экспрессируют аномальные рецепторы, которые постоянно находятся в активированном состоянии. Это приводит к патологическому росту клеток в ответ на физиологическую стимуляцию фактором роста или даже в отсутствие стимуляции фактором роста (лиганд-независимая передача сигналов).
Рецептор эпидермального фактора роста (epidermal growth factor receptor) часто избыточно экспрессируется в опухолях легких и желудочно-кишечного тракта, а HER2/neu — в злокачественных новообразованиях молочной железы. Оба рецептора активируют путь киназы Ras-Raf-митоген-активируемого белка (mitogen-activated protein), вызывая пролиферацию клеток.
4. Уклонение от супрессоров роста. В здоровых тканях межклеточный контакт в плотных клеточных популяциях действует как фактор, сдерживающий пролиферацию. Такое контактное торможение обычно отсутствует во многих тканях злокачественных новообразований. Факторы, ингибирующие рост, могут модулировать регуляторы клеточного цикла и активировать ингибиторы циклинзависимых киназ, вызывая ингибирование циклин-зависимых киназ. Мутации в ингибирующих белках часто обнаруживаются в злокачественных новообразованиях. Утрата рестрикции за счет нарушения регуляции pRB, обнаруживаемая в опухолях, приводит к нарушению сдерживания при переходе от фазы G к фазе S клеточного цикла.
Нарушение функции ТР53 оказывает последующее влияние на р21, что изменяет координацию репарации ДНК с остановкой цикла и приводит к накоплению геномных дефектов в пораженной клетке. Так, снижение уровней р21 и р27, которое можно обнаружить в опухолях с нормальной функцией ТР53, коррелирует с высокой степенью злокачественности опухоли и неблагоприятным прогнозом.
5. Включение репликативного бессмертия. Для развития злокачественных клеток в макроскопические опухоли необходима способность к неограниченному числу делений. Теломерные последовательности ДНК, которые защищают и стабилизируют концевые участки хромосом, играют основную роль в обеспечении этого неограниченного репликативного потенциала. В ходе репликации нормальных клеток теломеры постепенно укорачиваются, так как небольшие фрагменты теломерной ДНК утрачиваются при последовательных циклах репликации. Считается, что этот процесс укорочения представляет собой митотические часы и в конечном итоге препятствует дальнейшему делению клетки.
Теломераза, одна из разновидностей полимераз, добавляет нуклеотиды к теломерам, что позволяет клеткам продолжать деление, и таким образом предотвращает преждевременное прекращение клеточной репликации. Фермент теломераза почти полностью отсутствует в нормальных клетках, но экспрессируется в значительном количестве во многих злокачественных опухолевых клетках.
6. Стимуляция ангиогенеза. Все злокачественные новообразования нуждаются в развитой сосудистой сети для обеспечения постоянного роста. Без стимуляции развития новых сосудов опухоли не смогут вырасти более чем на 1 мм3. Новообразованиям жизненно необходимы питательные вещества и кислород, а также возможность избавляться от продуктов метаболизма и углекислого газа. Это влечет за собой образование новых кровеносных сосудов, которое называется ангиогенезом (рис. 2, 3).
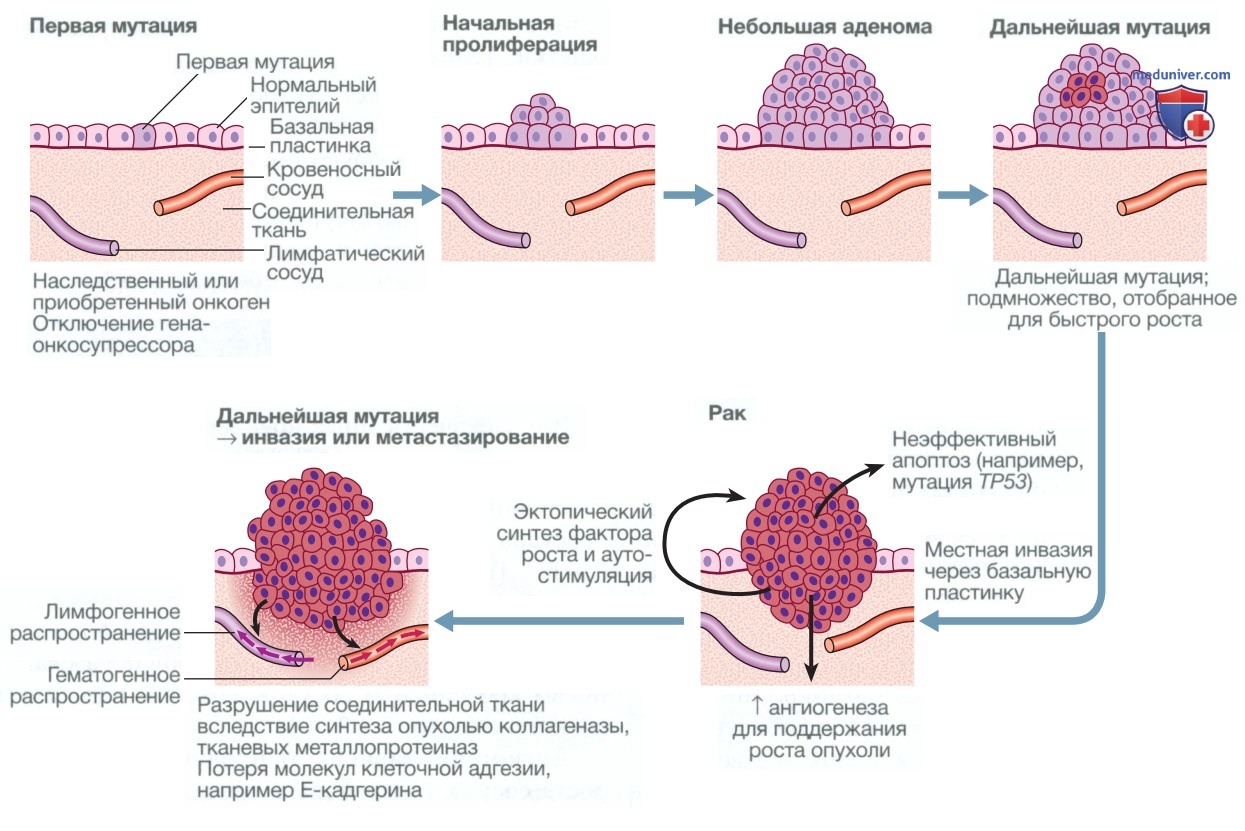
Рисунок 2. Онкогенез. Многоэтапное развитие злокачественной опухоли, включая инициацию опухоли, ее прогрессирование, инвазивный рост и метастазирование
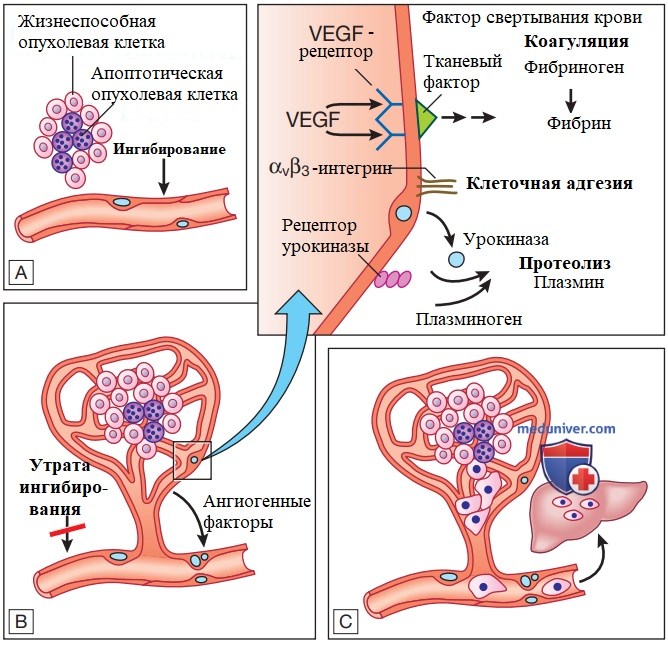
Рисунок 3. Ангиогенез, инвазия и метастазирование. А — для любого злокачественного новообразования, размерами превышающего 1 мм3, необходимо усиление кровоснабжения. В — образование новых сосудов является результатом высвобождения ангиогенных факторов опухолевыми клетками и утраты ингибирования эндотелиальными клетками. С — утрата клеточной адгезии и разрушение внеклеточного матрикса позволяют клеткам осуществлять экстравазацию в кровоток и метастазировать в отдаленные участки. VEGF — сосудистый эндотелиальный фактор роста
Ангиогенез зависит от продукции факторов роста сосудов, из которых лучше всего описаны сосудистый эндотелиальный фактор роста (vascular endothelial growth factor) и тромбоцитарный фактор роста. В процессе опухолевого роста активируется и поддерживается на интенсивном уровне ангиогенез, что способствует постоянному образованию сосудистой сетью новых сосудов, которые поддерживают прогрессию опухоли. Ангиогенез регулируется балансом проангиогенных стимулов и ингибиторов ангиогенеза. К последним относится тромбоспондин-1 (thrombospondin-1), который связывается с трансмембранными рецепторами эндотелиальных клеток и передает подавляющие сигналы.
Ряд клеток может участвовать в поддержании функциональной сосудистой сети опухоли и, следовательно, способствовать ангиогенезу. К ним относятся перициты и различные клетки костномозгового происхождения, такие как макрофаги, нейтрофилы, тучные клетки и миелоидные клетки-предшественники.
7. Активация инвазивного роста и метастазирование. Инвазивный рост и метастазирование представляют собой сложные процессы, включающие несколько отдельных этапов. Они начинаются с локальной тканевой инвазии с последующей инфильтрацией близлежащих кровеносных и лимфатических сосудов раковыми клетками. Затем злокачественные клетки гематогенным и лимфатическим путем переносятся в отдаленные участки организма, где образуют микрометастазы, которые в конечном итоге перерастают в макроскопические метастатические очаги (см. рис. 2).
Кадгерин-1 (cadherin-1) является кальций-зависимым гликопротеином клеточной адгезии, который облегчает сборку организованных клеточных пластов в тканях, а повышенная экспрессия считается антагонистом инвазивного роста и метастазирования. Опухоли in situ обычно продолжают продуцировать кадгерин-1, тогда как в злокачественных новообразованиях наблюдается прекращение продукции кадгерина-1 вследствие снижения синтеза или его инактивации под влиянием случайной мутации. Это подтверждает теорию, что кадгерин-1 играет ключевую роль в подавлении инвазивного роста опухоли и ее метастазирования.
Перекрестное взаимодействие между злокачественными клетками и клетками окружающей стромы приводит к появлению способности к инвазивному росту и метастазированию. Было выяснено, что мезенхимальные стволовые клетки в строме опухоли секретируют CCL5 — белок-хемокин, помогающий рекрутировать лейкоциты в очаги воспаления. С помощью ряда Т-клеточных цитокинов (ИЛ-2 и ИФН-γ) CCL5 индуцирует пролиферацию и активацию NK и затем реципрокно воздействует на раковые клетки, стимулируя их инвазивный рост. Макрофаги на периферии опухоли могут способствовать локальной инвазии, выделяя ферменты, разрушающие матрикс, такие как металлопротеиназы и цистеиновые протеазы (катепсины).
8. Перепрограммирование энергетического метаболизма. В аэробных условиях окислительное фосфорилирование функционирует как основной метаболический путь производства энергии. В ходе гликолиза клетки перерабатывают глюкозу сначала в пируват, а затем в углекислый газ в митохондриях. В анаэробных условиях гликолиз становится предпочтительным для получения аденозинтрифосфата. Раковые клетки могут перепрограммировать свой метаболизм глюкозы, ограничивая выработку энергии с помощью гликолиза даже в присутствии кислорода. Этот процесс получил название аэробного гликолиза. Регулирование с помощью переносчиков глюкозы, таких как GLUT1, является основным механизмом, посредством которого осуществляется аэробный гликолиз.
Такое перепрограммирование энергетического метаболизма представляется парадоксальным, поскольку общее производство энергии в результате гликолиза в 18 раз ниже, чем в результате окислительного фосфорилирования. Одним из объяснений может быть то, что повышенное образование промежуточных продуктов гликолиза помогает задействовать различные биосинтетические пути, в том числе такие, которые создают нуклеозиды и аминокислоты, необходимые для образования новых клеток.
9. Стимуляция воспаления опухолью. Почти во всех опухолевых тканях при патогистологическом исследовании обнаруживается инфильтрация иммунными клетками. Ранее считалось, что эти находки объясняются попыткой иммунной системы уничтожить раковую опухоль. В настоящее время стало очевидным, что связанные с опухолью воспалительные реакции способствуют образованию и прогрессированию рака.
Цитокины способны менять проницаемость кровеносных сосудов и обеспечивать миграцию лейкоцитов (главным образом нейтрофилов) из кровотока в ткани — этот процесс называется экс-травазацией. Миграция через эндотелий происходит посредством диапедеза, при котором градиенты хемокинов стимулируют адгезированные лейкоциты к перемещению между эндотелиальными клетками и прохождению через базальную мембрану в окружающие ткани. Оказавшись внутри тканевого интерстиция, лейкоциты связываются с белками внеклеточного матрикса посредством интегринов и CD44, предотвращая их исчезновение в данной области.
Наряду с клеточными медиаторами ряд бесклеточных биохимических каскадных систем, состоящих из предварительно образованных белков плазмы, действуют параллельно, инициируя и усиливая воспалительный ответ. К ним относятся система комплемента, активируемая бактериями, а также коагуляционная и фибринолитическая системы, активируемые некрозом, ожогами, травмами и самими злокачественными новообразованиями. Другие биологически активные молекулы, такие как факторы роста и проангиогенные факторы, могут высвобождать клетки зоны воспаления в микроокружение опухоли. В частности, высвобождение активных форм кислорода, которые являются активными мутагенами, будет ускорять генетическую эволюцию клеток, окружающих опухоль, стимулируя ее рост и способствуя прогрессированию рака.
10. Уклонение от иммунного ответа. Иммунная система служит важным барьером для образования и прогрессирования опухоли, а способность к уклонению от иммунного ответа является отличительной чертой развития злокачественных новообразований. Злокачественные клетки непрерывно вбрасывают поверхностные антигены в кровеносное русло, вызывая иммунный ответ, который включает продукцию цитотоксических Т-клеток, NK и макрофагов. Считается, что иммунная система обеспечивает непрерывный надзор с последующей элиминацией клеток, которые подвергаются злокачественной трансформации.
Однако нарушения в развитии или функционировании CD8+ цитотоксических Т-лимфоцитов, CD4+ Th1-хелперных Т-клеток и NK могут приводить к заметному увеличению вероятности развития рака. Высокоиммуногенные раковые клетки также могут уклоняться от иммунного надзора с помощью отключения компонентов иммунной системы. Это достигается за счет рекрутирования воспалительных клеток, включая регуляторные Т-клетки и миелоидные клетки-супрессоры, которые активно подавляют функционирование цитотоксических лимфоцитов.
Злокачественные новообразования развиваются и прогрессируют при нарушении их распознавания иммунной системой, снижении восприимчивости из-за уклонения от иммунного надзора и развитии дисфункции иммунной системы, зачастую посредством медиаторов воспаления.