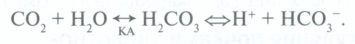Кислотно-щелочное равновесие (КЩР) - кратко с точки зрения внутренних болезней
Значение pH артериальной плазмы обычно составляет 7,40, что соответствует концентрации Н+ 40 нмоль/л, и в норме концентрации Н+ не выходят за пределы диапазона 37—45 нмоль/л (pH 7,43—7,35). Нарушения кислотно-щелочного равновесия (КЩР) могут возникать при самых разных заболеваниях. Увеличение концентрации Н+ вызывает ацидоз с уменьшением pH, тогда как снижение концентрации Н+ вызывает алкалоз с повышением pH.
а) Функциональная анатомия и физиология. Разнообразные физиологические механизмы поддерживают pH ВКЖ в узком диапазоне. Сначала действуют буферы крови и тканей; наиболее важным является реакция ионов Н+ с гидрокарбонатом с образованием углекислоты, которая под действием фермента карбоангидразы (КА) диссоциирует с образованием СО2 и воды:
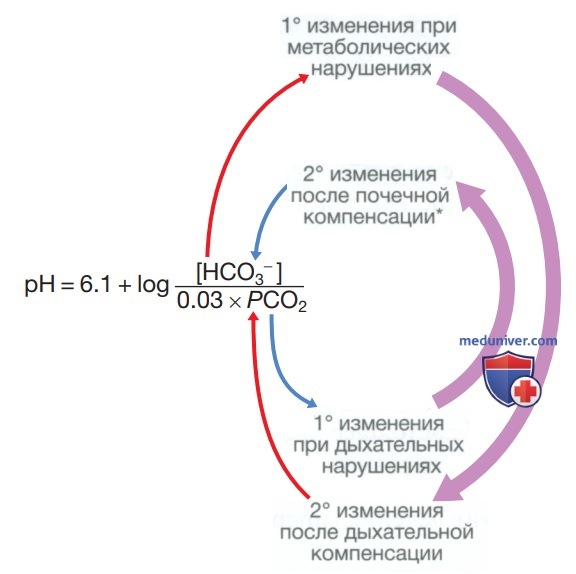
Эта буферная система важна, поскольку концентрация гидрокарбоната в ВКЖ относительно высока (21—29 ммоль/л), и два его основных компонента находятся под физиологическим контролем: СО2 (легкие) и гидрокарбонат (почки). Эти взаимосвязи проиллюстрированы на рис. ниже (форма уравнения Хендерсона-Хассельбальха).
Взаимосвязь между pH, рCO2 (в мм рт.ст.) и концентрацией гидрокарбоната в плазме крови (в ммоль/л). Следует отметить, что изменения концентрации НСО3- также являются частью почечной коррекции при устойчивых метаболических отклонениях кислотно-щелочного равновесия до тех пор, пока сами почки не станут причиной первичного нарушения
При нарушениях КЩР возможна быстрая дыхательная компенсация. В ответ на накопление кислоты изменения pH в стволе головного мозга стимулируют дыхание, снижают рСО2 и повышают pH. И наоборот, системный алкалоз приводит к угнетению дыхания, вызывая повышение рСO2 и снижение pH, хотя следует отметить, что этот механизм имеет ограниченную способность изменять pH, поскольку гипоксия представляет собой альтернативный стимул для управления дыханием.
Почки обеспечивают третью линию защиты от нарушений pH артериальной крови. Когда кислота накапливается из-за хронических дыхательных или метаболических (внепочечных) причин, почки в течение длительного времени способны усиливать экскрецию кислоты с мочой, эффективно увеличивая содержание гидрокарбоната в плазме крови.
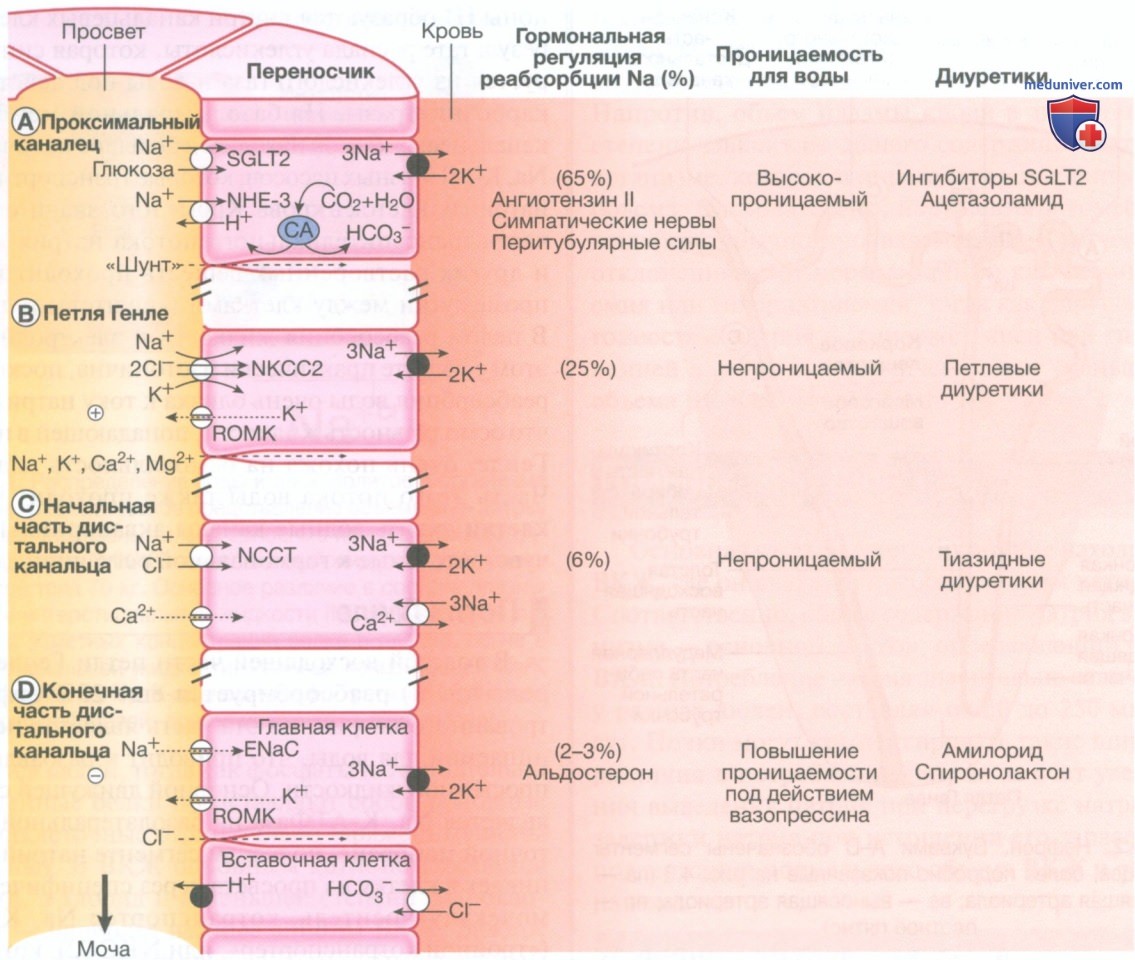
- Регуляция почками кислотнощелочного равновесия. Регуляция КЩР происходит на нескольких участках почек. В проксимальных канальцах реабсорбируется около 85% отфильтрованных ионов гидрокарбоната с помощью механизма секреции Н+, показанного на рис. ниже, А. Этот процесс зависит от фермента карбоангидразы, как в цитоплазме клеток проксимальных канальцев, так и на люминальной поверхности мембран щеточной каймы. Система обладает высокой емкостью и необходима для возвращения отфильтрованного гидрокарбоната, но не приводит к значительному закислению просветной жидкости.
Основные транспортные механизмы в сегментах нефрона. Апикальная мембрана канальцевых клеток — это сторона, обращенная в просвет, а базолатеральная мембрана — это сторона, обращенная в кровеносный сосуд. Черными кружками обозначены активные транспортные насосы, связанные с гидролизом аденозинтрифосфата, а белые символы обозначают ионные каналы и молекулы-переносчики. Показана доля реабсорбированного натрия, влияние регуляторных факторов, проницаемость для воды и места действия различных классов диуретиков. Ингибиторы натрий-зависимого транспортера глюкозы 2 в основном используются при лечении сахарного диабета, но обладают мочегонными свойствами, блокируя натрий-зависимый транспортер глюкозы 2 в проксимальных канальцах. В том же месте ацетазоламид ингибирует карбоангидразу (КА), которая, уменьшая выработку ионов водорода проксимальными канальцевыми клетками, ингибирует обмен натрия и водорода с помощью переносчика NHE-3. Петлевые диуретики блокируют переносчик NKCC2 в петле Генле, тогда как тиазидные диуретики ингибируют канал котранспортера натрия и хлорида в начальной части дистального канальца. Амилорид и спиронолактон блокируют эпителиальный натриевый канал в конечной части дистального канальца и собирающих трубочках.
Дистальные сегменты нефрона также играют важную роль в экскреции кислот. Ионы водорода секретируются в просвет с помощью Н+-АТФазы во вставочных клетках кортикального отдела и во внешних клетках медуллярного отдела собирательной трубочки. Ионы Н+ образуются в клетках канальцев в результате гидратации СO2 с образованием углекислоты, которая диссоциирует на ион Н+, выделяющийся в просвет канальца, и ион гидрокарбоната, который проходит через базолатеральную мембрану в кровь.
Секретируемые ионы Н+ способствуют реабсорбции остаточного гидрокарбоната из просветной жидкости за счет образования внутриклеточного иона ОН-, реагирующего с СO2 с образованием НСO3-, который выходит через базолатеральную мембрану. Тем не менее секреция Н+ также способствует выведению из организма кислот, связанных с различными мочевыми буферами, из которых наиболее важными являются фосфат и аммиак. Мочевые буферы необходимы для предотвращения снижения pH мочи, что может создать неблагоприятный градиент, который может блокировать дальнейшую секрецию Н+.
Отфильтрованный фосфат (Н2РО42-) объединяется с Н+ в просвете дистального канальца с образованием ди гидрофосфата (Н2РО42-), который выделяется в мочу с натрием. Аммиак (NH3) образуется в канальцевых клетках при дезаминировании аминокислоты глутамина ферментом глутаминазой. Затем NH3 реагирует с секретируемым Н+в просвете канальца с образованием иона аммония (NH4+), который попадает в просветную жидкость и выделяется с ионами хлоридов.
Эти два механизма выводят из организма примерно 1 ммоль/кг ионов водорода в сутки, что соответствует нелетучей кислотной нагрузке в результате метаболизма пищевых белков. Слабощелочное значение pH плазмы крови 7,4 (Н+ — 40 нмоль/л), которое поддерживается в здоровом организме, можно объяснить способностью почек вырабатывать кислую мочу (обычно pH 5—6 (Н+ — 1 000— 10 000 нмоль/л), в которую может выделяться чистый суточный избыток метаболических кислот, вырабатываемых организмом.
б) Клинические проявления нарушений кислотно-щелочного равновесия. Отклонения КЩР могут клинически проявляться либо нарушением работы тканей из-за изменения pH (например, дисфункция сердечной мышцы или центральной нервной системы), либо вторичными изменениями дыхания, которые возникают как реакция на основное метаболическое изменение (например, дыхание Куссмауля при метаболическом ацидозе). В клинической картине часто преобладает основная причина, а не симптомы нарушения КЩР как таковые. Нарушения КЩР часто становятся очевидными только при отклонении от нормы концентрации гидрокарбоната в венозной плазме или изменениях pH, pCO2 или гидрокарбоната при анализе газового состава крови.
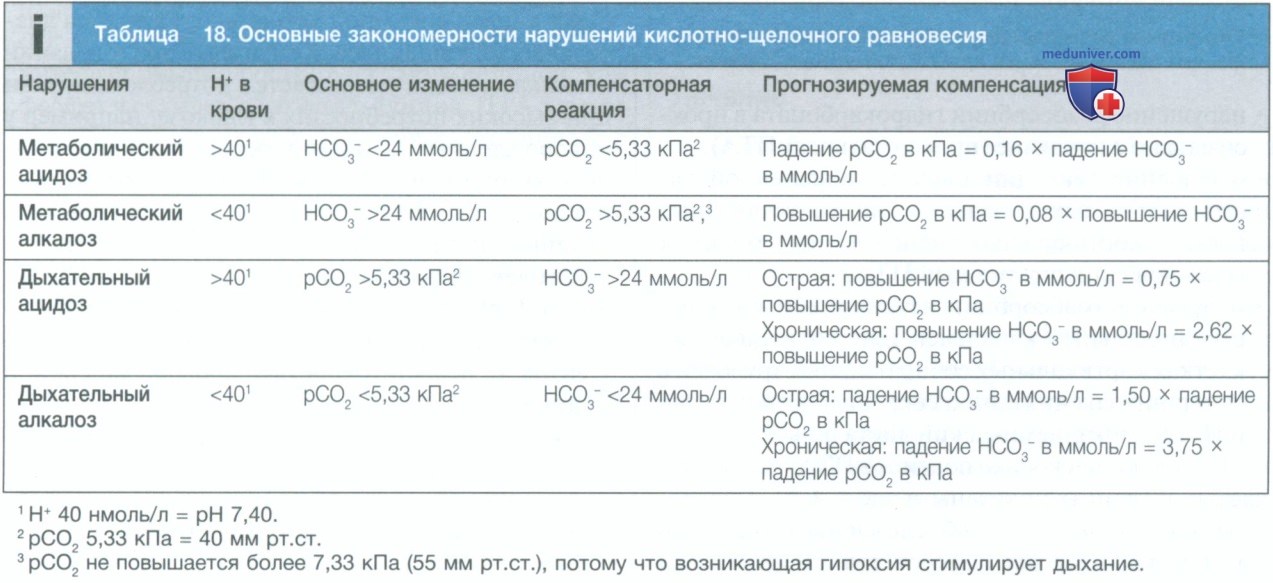
Наиболее частые закономерности при нарушениях газового состава крови показаны в табл. 18. Следует отметить, что термины «ацидоз» и «алкалоз» относятся исключительно к изменениям, лежащим в основе изменений КЩР, в то время как термины «ацидемия» и «алкалемия» более правильно относить только к характеристикам изменений в крови.
При метаболических нарушениях дыхательная компенсация происходит практически мгновенно, поэтому прогнозируемое компенсаторное изменение рСО2 достигается вскоре после развития метаболического нарушения. С другой стороны, при дыхательных нарушениях происходит небольшое первоначальное изменение уровня гидрокарбоната в результате химической буферизации CO2, в основном в эритроцитах, но через несколько дней или недель почки обеспечивают дальнейшие компенсаторные изменения концентрации гидрокарбоната в результате длительного изменения способности к секреции кислот.
Когда полученные клинически кислотно-основные параметры не соответствуют прогнозируемой компенсации, следует подозревать смешанное нарушение КЩР.
Видео урок по анализу КЩС в норме и его расшифровка