Функциональная анатомия и физиология крови - кратко с точки зрения внутренних болезней
Кровь движется по сосудистой системе и состоит:
• из эритроцитов, транспортирующих кислород из легких в ткани;
• лейкоцитов, защищающих организм от инфекций;
• тромбоцитов, взаимодействующих с кровеносными сосудами и факторами свертывания крови, для поддержания целостности сосудов и предотвращения кровотечений;
• плазмы, содержащей многофункциональные белки, к которым относят антитела и факторы свертывания крови.
а) Гемопоэз. Под термином «гемопоэз» понимают образование клеток крови. Это активный процесс, который должен поддерживать нормальный уровень циркулирующих клеток и быть способен быстро реагировать на повышение потребности в них, как, например, при кровотечении или инфекции. Во время развития эмбриона гемопоэз происходит в желточном мешке, печени и селезенке, а затем в красном костном мозге, находящемся в медуллярных полостях всех костей. В детском возрасте красный костный мозг постепенно замещается жиром (желтый костный мозг), поэтому у взрослых гемопоэз в норме происходит только в позвонках, костях таза, грудине, ребрах, ключицах, черепе, верхней части плечевых костей и проксимальной части бедренных костей. Тем не менее красный костный мозг может разрастаться в ответ на повышенную потребность организма в клетках крови.
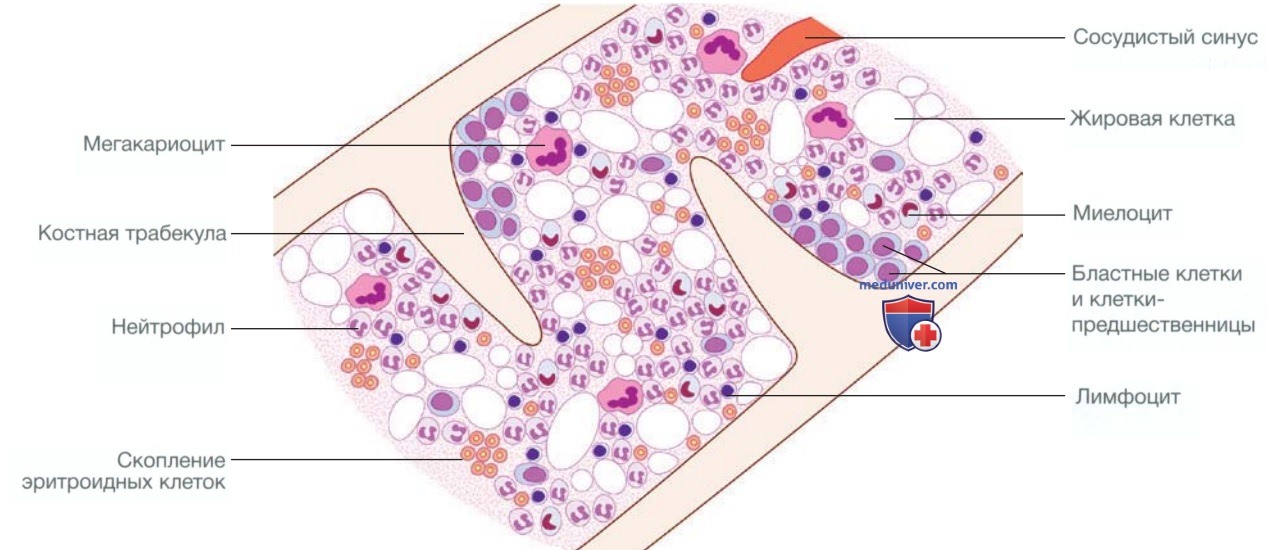
Костный мозг содержит ряд незрелых гемопоэтических клеток-предшественников и пул зрелых клеток для высвобождения в периоды повышенной потребности. Гемопоэтические клетки тесно взаимодействуют с окружающей стромой соединительной ткани, состоящей из ретикулярных клеток, макрофагов, жировых клеток, кровеносных сосудов и нервных волокон (рис. 1). В норме в костном мозге скопления клеток-предшественников эритроцитов сгруппированы вокруг центрального макрофага, который является источником железа и фагоцитирует ядра эритроцитов перед их выходом в кровоток. Мегакариоциты — это крупные клетки, которые образуют и высвобождают тромбоциты в сосудистые синусы. Клетки-предшественники лейкоцитов сгруппированы рядом с костными трабекулами; созревающие клетки мигрируют в пространства костного мозга к сосудистым синусам.
Рисунок 1. Структура костного мозга в норме
Плазматические клетки представляют собой зрелые В-клетки, секретирующие антитела, которые обычно составляют менее 5% популяции клеток костного мозга и разбросаны по межтрабекулярным пространствам.
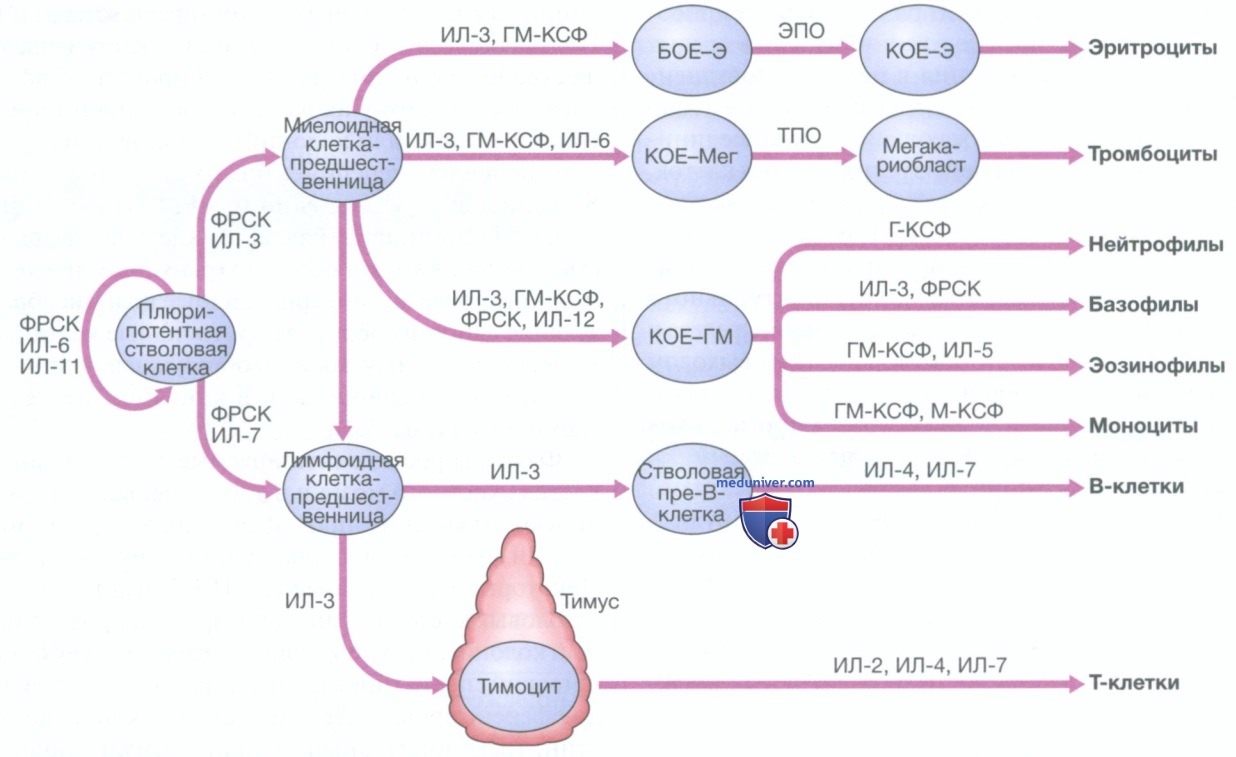
1. Стволовые клетки. Все клетки крови развиваются из плюрипотентных гемопоэтических стволовых клеток. Они составляют лишь 0,01% от общего количества клеток костного мозга, но могут самообновляться (т.е. образовывать большее число стволовых клеток) или дифференцироваться, создавая иерархию линии коммитированных клеток-предшественниц. Получающиеся в результате незрелые клетки-предшественницы нельзя идентифицировать морфологически, поэтому их называют в соответствии с типами клеток (или колоний), которые они образуют во время экспериментов в культурах клеток. Колониеобразующая единица-ГМ (гранулоцитарная, моноцитарная) является клеткой-предшественницей, которая образует гранулоцитарные и моноцитарные линии, эритроидные колониеобразующие единицы образуют эритроидные клетки, а мегакариоцитарная колониеобразующая единица образует мегакариоциты и в конечном итоге — тромбоциты (рис. 2).
Рисунок 2. Стволовые клетки и факторы роста в процессе развития гемопоэтических клеток: БОЕ-Э — бурстообразующая единица — эритроидная; КОЕ-Э — колониеобразующая единица—эритроидная; КОЕ-ГМ — колониеобразующая единица — гранулоцитарная, моноцитарная; КОЕ-Мег — колониеобразующая единица — мегакариоцитарная; ЭПО — эритропоэтин; Г-КСФ — гранулоцитарный колониестимулирующий фактор; ГМ-КСФ — гранулоцитарно-макрофагальный колониестимулирующий фактор; М-КСФ — макрофагальный колониестимулирующий фактор; ФРСК — фактор роста стволовых клеток; ТПО — тромбопоэтин
Факторы роста, синтезируемые в стромальных клетках костного мозга и в других местах, контролируют выживание, пролиферацию, дифференцировку и функцию стволовых клеток и их потомков. Некоторые из них, такие как ИЛ-3, фактор роста стволовых клеток и гранулоцитарно-макрофагальный колониестимулирующий фактор, действуют на широкий спектр типов клеток на различных стадиях дифференцировки. Другие, такие как эритропоэтин, гранулоцитарный колониестимулирующий фактор и тромбопоэтин, являются специфичными для клеточной линии. Многие из этих факторов роста в настоящее время синтезируются с помощью технологии рекомбинантной ДНК и используются в качестве средств терапии: например, эритропоэтин для лечения анемии на фоне почечной недостаточности и гранулоцитарного колониестимулирующего фактора для ускорения восстановления нейтрофилов после химиотерапии.
Костный мозг также содержит стволовые клетки, которые могут дифференцироваться в негематологические клетки. Мезенхимальные стволовые клетки дифференцируются в клетки скелетных мышц, хрящевые клетки, клетки сердечной мышцы и жировые клетки, в то время как другие стволовые клетки дифференцируются в клетки нервов, печени и эндотелия кровеносных сосудов. Это называется пластичностью стволовых клеток, и в будущем этот феномен может иметь многообещающее клиническое применение.
б) Клетки крови и их функции:
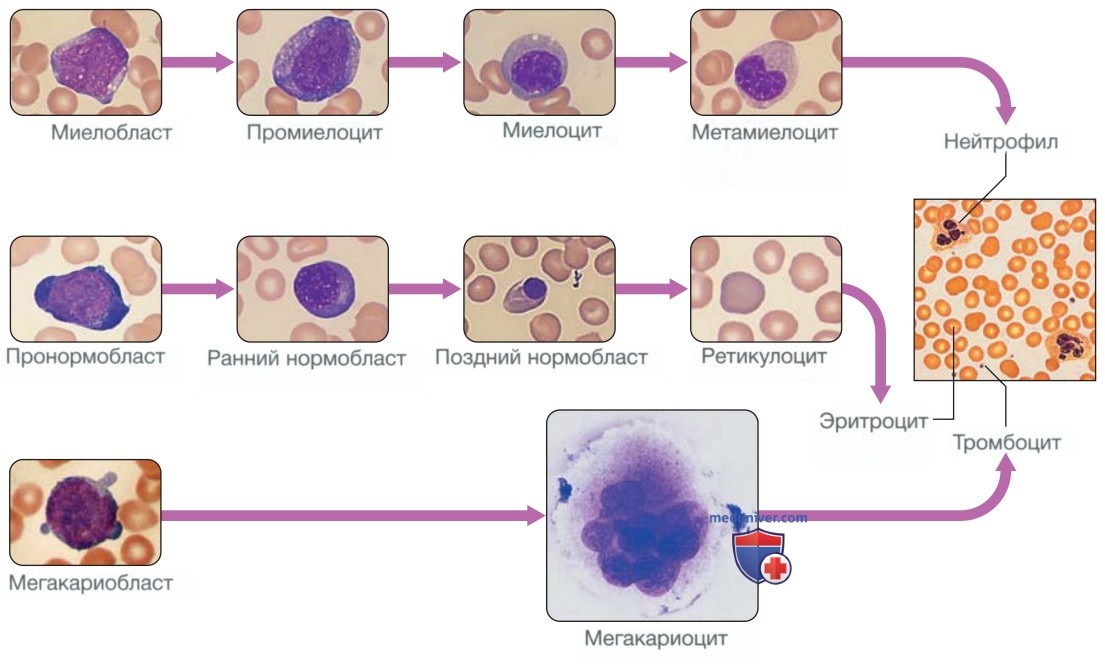
1. Эритроциты. Предшественники эритроцитов, образовавшиеся в костном мозге из эритроидных клеток-предшественниц (эритроидных колониеобразующих единиц), называются эритробластами или нормобластами (рис. 3). Ониделятся и приобретают гемоглобин, который окрашивает цитоплазму в розовый цвет; ядро конденсируется и выталкивается из клетки. Первый безъядерный эритроцит — это ретикулоцит, который все еще содержит в цитоплазме рибосомный материал, придающий этим крупным клеткам синеватый оттенок (полихромазия). Ретикулоциты утрачивают свой рибосомный материал и созревают в течение трех дней, когда они попадают в кровоток. Увеличение количества циркулирующих ретикулоцитов (ретикулоцитоз) отражает усиление эритропоэза.
Рисунок 3. Путь созревания эритроцитов, гранулоцитов и тромбоцитов. Справа показан мазок крови в норме
Пролиферация и дифференцировка клеток-предшественниц эритроцитов стимулируется эритропоэтином, полипептидным гормоном, синтезируемым почечными интерстициальными перитубулярными клетками в ответ на гипоксию. Нарушение синтеза эритропоэтина у пациентов с почечной недостаточностью вызывает анемию, которую можно лечить экзогенным рекомбинантным эритропоэтином или аналогичными ЛС, стимулирующими эритропоэз (например, дарбэпоэтином альфа).
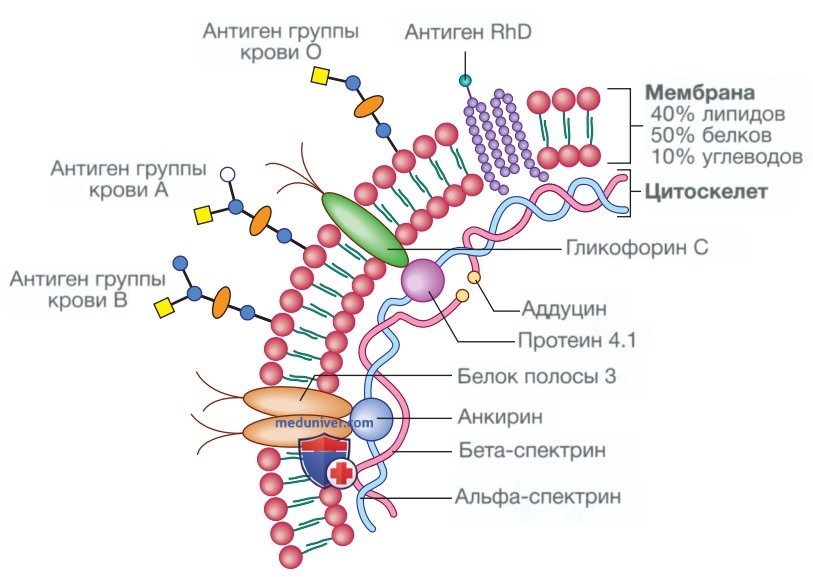
В норме зрелые эритроциты циркулируют в крови около 120 дней. Это двояковогнутые диски диаметром 8 мкм, без ядра, но заполненные гемоглобином, который доставляет кислород к тканям. Чтобы пройти через самые маленькие капилляры, мембрана эритроцитов способна деформироваться благодаря липидному бислою, к которому с помощью специальных связывающих белков прикреплен «скелет» из филаментных белков (рис. 4). Наследственные патологии любого из этих белков приводят к потере мембраны при прохождении клеток через селезенку и образованию эритроцитов неправильной формы, называемых сфероцитами, или овалоцитами (см. рис. 5.8).
Рисунок 4. Структура мембраны эритроцитов в норме. Гибкость эритроцитарной мембраны обеспечивается благодаря прикреплению белков цитоскелета. Важные трансмембранные белки включают белок полосы 3 (ионный транспортный канал) и гликофорин С (участвует в прикреплении цитоскелета и газообмене, а также является рецептором для Plasmodium falciparum при малярии). Антигены в эритроцитах определяют группу крови человека. Существует около 22 систем групп крови (группы углеводных или белковых антигенов, контролируемых одним геном или несколькими тесно связанными локусами); клинически наиболее важными являются системы АВ0 и резус-фактор (Rh). Генетический локус АВ0 имеет три основные аллельные формы: А, В и 0. Аллели А и В кодируют гликозилтрансферазы, которые вводят N-ацетилгалакто-замин (белый круг) и D-галактозу (синий круг) соответственно в молекулы углеводных антигенов на поверхности мембраны. У людей с аллелем 0 синтезируется антиген 0, в котором отсутствуют какие-либо из этих добавленных групп сахаров. Резус-антигены — это трансмембранные белки
Эритроциты подвергаются осмотическому стрессу в легочном и почечном кровотоке; для поддержания гомеостаза мембрана содержит ионные насосы, которые контролируют внутриклеточные уровни натрия, калия, хлорида и гидрокарбоната. При отсутствии митохондрий энергия для этих функций поставляется за счет анаэробного гликолиза и пентозофосфатного пути в цитозоле. Мембранные гликопротеины в липидном бислое также образуют антигены, распознаваемые по группам крови (см. рис. 4). Системы АВО и резус-фактор являются наиболее распространенными, но было описано более 400 антигенов групп крови.
- Гемоглобин. Гемоглобин — это белок, специально адаптированный для транспорта кислорода. Он состоит из четырех цепей глобина, каждая из которых окружает железосодержащую порфириновую пигментную молекулу, называемую гемом. Глобиновые цепи представляют собой комбинацию двух альфа-цепей и двух не-альфа-цепей; гемоглобин А (αα/ββ) составляет более 90% гемоглобина у взрослых, тогда как гемоглобин F (αα/γγ) является преобладающим типом гемоглобина у плода. Каждая молекула гема содержит ион железа (Fe2+), с которым обратимо связывается кислород; сродство к кислороду возрастает по мере связывания последующих молекул кислорода. Когда кислород связан, бета-цепи «сходятся» вместе; после высвобождения кислорода они расходятся.
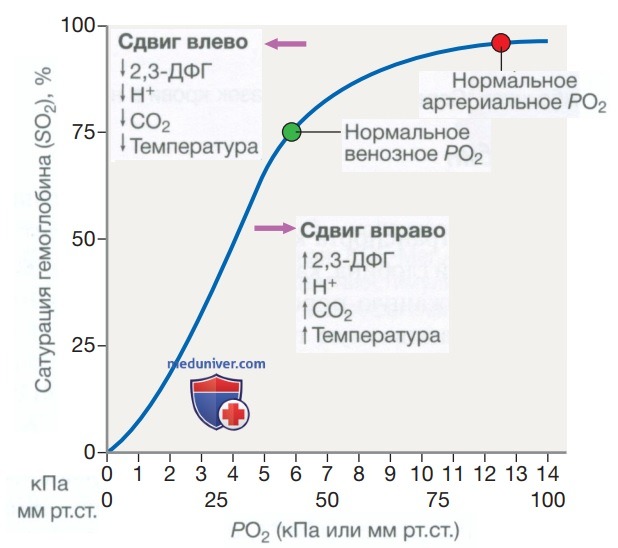
В «открытом» дезоксигенированном состоянии 2,3-дифосфоглицерат, продукт метаболизма эритроцитов, связывается с молекулой гемоглобина и снижает ее сродство к кислороду. Эти сложные взаимодействия обусловливают сигмовидную форму кривой диссоциации кислорода (рис. 5). Положение этой кривой зависит от концентрации 2,3-дифосфоглицерата, ионов Н+ и CO2; повышение их уровней сдвигает кривую вправо и облегчает выделение кислорода, например когда эритроциты достигают гипоксических тканей. Гемоглобин F не способен связывать 2,3-дифосфоглицерат и характеризуется смещением влево кривой диссоциации кислорода, что наряду с низким значением pH фетальной крови обеспечивает оксигенацию плода. Сильные окислители, такие как дапсон, могут превращать двухвалентное железо в гемоглобине в его трехвалентное состояние (Fe3+). Образующийся в результате метгемоглобин также имеет смещенную влево кривую диссоциации кислорода, что может привести к гипоксии тканей.
Рисунок 5. Кривая диссоциации гемоглобина и кислорода. Перечислены факторы, которые смещают кривую вправо (из крови выделяется больше кислорода) и влево (выделяется меньше кислорода) при определенном значении PO2. Чтобы преобразовать кПа в мм рт.ст., умножьте на 7,5 (2,3-ДФГ — 2,3-дифосфоглицерат)
Генетические мутации, изменяющие гемсвязывающие карманы цепей глобина или «шарнирные» взаимодействия между цепями глобина, приводят к развитию гемоглобинопатий или образованию нестабильного гемоглобина. Альфа-глобиновые цепи кодируются двумя генами в хромосоме 16, а бета-глобиновые цепи — одним геном в хромосоме 11; дисбаланс в образовании цепей глобина приводит к талассемии. Нарушения в синтезе гема вызывают порфирии.
- Разрушение эритроцитов. В конце своей жизни продолжительностью приблизительно 120 дней эритроциты фагоцитируются ретикулоэндотелиальной системой. Аминокислоты из цепей глобина возвращаются в метаболизм, а железо извлекается из гема для повторного использования в синтезе гемоглобина. Остатки гемовой структуры разлагаются до билирубина, конъюгируют с глюкуроновой кислотой и выводятся с желчью. В тонкой кишке билирубин превращается в стеркобилин; большая часть стеркобилина выводится из организма, но небольшое количество реабсорбируется и выводится почками в виде уробилиногена. Усиленное разрушение эритроцитов из-за гемолиза или неэффективного гемопоэза приводит к развитию желтухи и увеличению содержания уробилиногена в моче. Свободный внутрисосудистый гемоглобин токсичен и обычно связан с гаптоглобинами — белками плазмы крови, образующимися в печени.
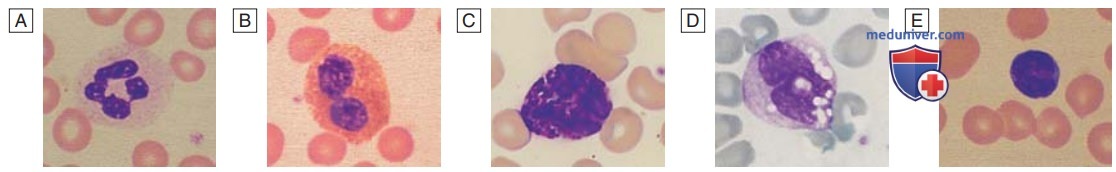
2. Лейкоциты. Лейкоциты, или белые кровяные клетки, состоят из гранулоцитов (нейтрофилов, эозинофилов и базофилов), моноцитов и лимфоцитов (см. рис. ниже). Гранулоциты и моноциты образуются из клеток-предшественниц гранулоцитарных, моноцитарных колониеобразующих единиц при миелопоэзе в костном мозге. Первым распознаваемым гранулоцитом в костном мозге является миелобласт — крупная клетка с небольшим количеством базофильной цитоплазмы и примитивным ядром с открытым хроматином и ядрышками. По мере деления и созревания клеток сегменты ядра и цитоплазмы приобретают специфические нейтрофильные, эозинофильные или базофильные гранулы (см. рис. 3).
Внешний вид лейкоцитов: А — нейтрофил; B — эозинофил; C — базофил; D — моноцит; E — лимфоцит
Этот процесс занимает около 14 дней. Цитокины гранулоцитарного колониестимулирующего фактора, гранулоцитарно-макрофагального колониестимулирующего фактора и М-колониестимулирующего фактора участвуют в образовании миелоидных клеток, а гранулоцитарный колониестимулирующий фактор можно использовать в клинической практике и для ускорения восстановления количества нейтрофилов в крови после химиотерапии.
Миелоциты или метамиелоциты обычно находятся только в костном мозге, но могут появляться в кровотоке при инфекциях или интоксикациях. Появление более ранних миелоидных предшественников в крови часто сопровождается наличием ядросодержащих эритроцитов и называется лейкоэритробластной картиной; это указывает на серьезное нарушение функции костного мозга.
- Нейтрофилы. Нейтрофилы — это наиболее многочисленная группа лейкоцитов в крови у взрослых. Они имеют диаметр 10—14 мкм, с дольчатым ядром, содержащим 2—5 сегментов, и гранулами в цитоплазме. Их основная функция — распознавать, поглощать и уничтожать чужеродные частицы и микроорганизмы. В костном мозге находится большой пул зрелых нейтрофилов. Каждый день около 1011 нейтрофилов попадают в кровоток, где клетки могут свободно циркулировать и прикрепляться к эндотелию по периферии участка воспаления. Эти два пула сопоставимы по размеру; такие факторы, как физическая нагрузка и выброс катехоламинов, увеличивают количество клеток в крови. Нейтрофилы проводят в кровотоке 6—10 ч, а затем элиминируются главным образом селезенкой. Кроме того, они могут проникать в ткани и участвовать в воспалительном процессе или подвергаться апоптозу и быть фагоцитированы макрофагами.
- Эозинофилы. На долю эозинофилов приходится 1—6% общего числа циркулирующих лейкоцитов. Они похожи по размеру на нейтрофилы, но имеют двудольное ядро и заметные оранжевые гранулы при окрашивании по Романовскому. Эозинофилы способны к фагоцитозу, и их гранулы содержат пероксидазу, способную генерировать активные формы кислорода и белки, участвующие во внутриклеточном уничтожении простейших и гельминтов. Они также участвуют в аллергических реакциях (например, при атопической бронхиальной астме).
- Базофилы. Эти клетки встречаются реже, чем эозинофилы, составляя менее 1% общего числа циркулирующих лейкоцитов. Они содержат плотные черные гранулы, за которыми не видно ядра. Тучные клетки напоминают базофилы, но присутствуют только в тканях. Эти клетки участвуют в реакциях гиперчувствительности.
- Моноциты. Моноциты — самые крупные среди всех лейкоцитов клетки диаметром 12—20 мкм и ядром неправильной формы, находящемся в большом объеме бледно-голубой цитоплазмы, содержащей редкие цитоплазматические вакуоли. Эти клетки циркулируют в крови в течение нескольких часов, а затем мигрируют в ткани, где превращаются в макрофаги, клетки Купфера или антигенпрезентирующие дендритные клетки. Макрофаги фагоцитируют остатки клеток, апоптотические клетки и микроорганизмы.
- Лимфоциты. Лимфоциты происходят из плюрипотентных гемопоэтических стволовых клеток в костном мозге. Выделяют два основных типа лимфоцитов: Т-клетки (которые опосредуют клеточный иммунитет) и В-клетки (которые опосредуют гуморальный иммунитет). Лимфоидные клетки, которые мигрируют в тимус, развиваются в Т-клетки, тогда как В-клетки развиваются в костном мозге.
Большинство (около 80%) лимфоцитов в кровотоке — это Т-клетки. Лимфоциты неоднородны, самые маленькие по размеру похожи на эритроциты, а самые большие — на нейтрофилы. Маленькие лимфоциты имеют круглую форму со скудной цитоплазмой, но более крупные клетки характеризуются неправильной формой с большим объемом голубой цитоплазмы. У субпопуляций лимфоцитов есть определенные функции, а продолжительность их жизни может составлять от нескольких дней до нескольких лет. Антигены клеточной поверхности (антигены «кластера дифференцировки»), которые появляются в разные моменты созревания лимфоцитов и указывают на происхождение и зрелость клетки, используются для классификации лимфом и лимфолейкозов.
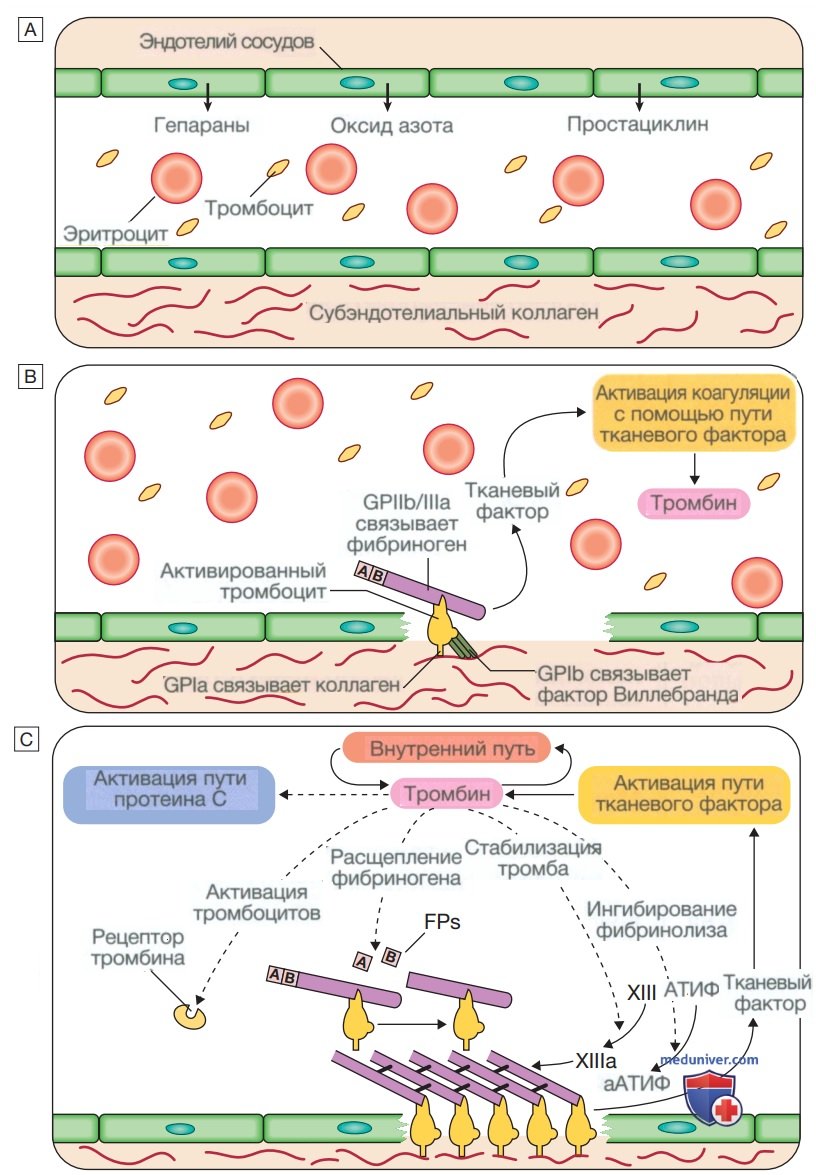
в) Гемостаз. Кровь должна поддерживаться в жидком состоянии, чтобы она могла функционировать в качестве транспортной системы, но она должна быть способной сворачиваться, образуя сгусток после повреждения сосудов, чтобы предотвратить чрезмерное кровотечение, — этот процесс называется гемостазом. Успешный гемостаз происходит в месте повреждения, и в дальнейшем происходят растворение сгустка и восстановление ткани. Это достигается за счет сложных взаимодействий между эндотелием сосудов, тромбоцитами, фактором Виллебранда (von Willebrand factor — vWF), факторами свертывания крови, естественными антикоагулянтами и фибринолитическими ферментами (рис. 6). Дисфункция любого из этих компонентов может привести к кровотечению или тромбозу.
Рисунок 6. Стадии гемостаза в норме. А — стадия 1. Условия перед травмой не мешают кровотоку. Эндотелий сосудов вырабатывает вещества (в том числе оксид азота, простациклин и гепариноподобные вещества) для предотвращения адгезии тромбоцитов и лейкоцитов к стенке сосуда. Тромбоциты и факторы свертывания крови циркулируют в неактивном состоянии. В — стадия 2. Ранний гемостатический ответ: тромбоциты адгезируются; активируется коагуляция. В месте повреждения сосуда эндотелий разрушается, обнажая субэндотелиальный коллаген. Выделяются небольшие количества тканевого фактора (ТФ). Тромбоциты связываются с коллагеном с помощью специфического рецептора гликопротеина la (GPIa), что вызывает изменение формы тромбоцитов и их адгезию к области повреждения за счет связывания других рецепторов (GPIb и GPIIb/llla) с фактором Виллебранда и фибриногеном соответственно. Коагуляция активируется тканевым фактором (внешний путь) с образованием небольшого количества тромбина. С и D — стадия 3. Формирование фибринового сгустка: тромбоциты активируются и агрегируют; образование фибрина поддерживается мембраной тромбоцитов: образуется стабильный фибриновый сгусток. Адгезирующиеся тромбоциты активируются многими путями, включая связывание аденозиндифосфата, коллагена, тромбина и адреналина (эпинефрина) с поверхностными рецепторами. Циклооксигеназный путь превращает арахидоновую кислоту из мембраны тромбоцитов в тромбоксан А2, который вызывает агрегацию тромбоцитов. Активация тромбоцитов приводит к высвобождению содержимого их гранул, что еще больше усиливает коагуляцию (см. рис. 7). Тромбин играет основную роль в контроле коагуляции: небольшое количество, образующееся через путь тканевого фактора, значительно усиливает его собственный синтез; «внутренний» путь становится активным с образованием большого количества тромбина. Тромбин непосредственно вызывает образование тромба за счет отщепления фибринопептидов (ФП) от фибриногена с образованием фибрина. Мономеры фибрина «сшиты» фактором XIII, который также активируется тромбином.
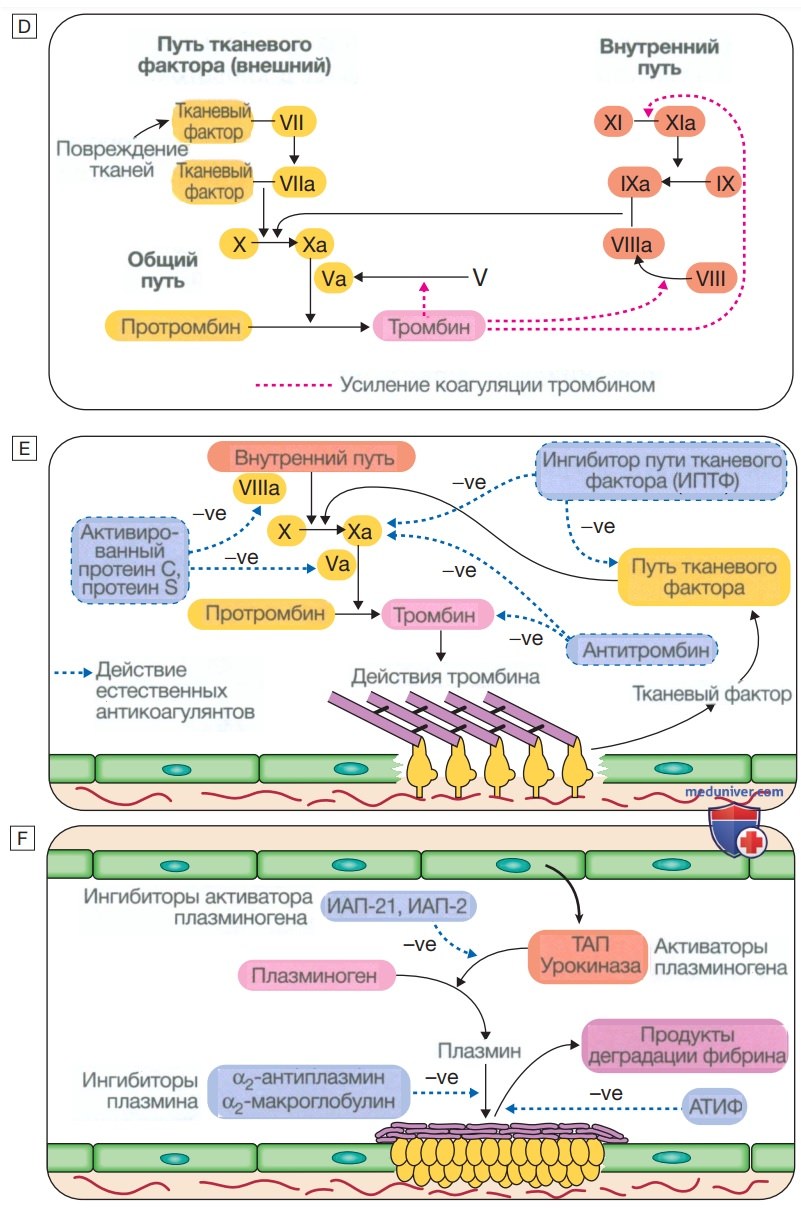
Рисунок 6. (Окончание) Сыграв ключевую роль в образовании и стабилизации тромба, тромбин начинает регулировать образование тромба двумя основными путями: (а) активация пути протеина С (PC) (естественный антикоагулянт), который уменьшает дальнейшую коагуляцию; (b) активация активируемого тромбином ингибитора фибринолиза (АТИФ), который ингибирует фибринолиз (см. Е и F). Д — стадия 4. Ограничение образования тромба: естественные антикоагулянты прекращают активацию факторов свертывания крови. После достижения гемостаза дальнейшее увеличение тромба ограничивается антикоагулянтами. Антитромбин — это ингибитор сериновой протеазы, синтезируемый в печени, который разрушает активированные факторы, такие как XIa, Ха и тромбин (IIа). Его основная активность против тромбина и Ха усиливается гепарином и фондапаринуксом натрия, что объясняет их антикоагулянтный эффект. Ингибитор пути тканевого фактора (ИПТФ) связывается с факторами Vila и Ха и инактивирует их. Активация протеина С происходит после связывания тромбина с мембраносвязанным тромбомодулином; активированный протеин С (аРС) связывается со своим кофактором, протеином S (PS), и расщепляет Va и VIIIa. Протеин С и протеин S зависят от витамина К, и их запасы истощаются кумариновыми антикоагулянтами, такими как варфарин. Е — стадия 5. Фибринолиз: плазмин разрушает фибрин, чтобы обеспечить реканализацию сосуда и восстановление тканей. Нерастворимый тромб необходимо разрушить для реканализации сосуда. Плазмин, основной фибринолитический фермент, образуется при активации плазминогена, например тканевым активатором плазминогена (ТАП) или урокиназой в тромбе. Плазмин гидролизует фибриновый тромб с образованием продуктов деградации фибрина, в том числе D-димера. Этот процесс строго регулируется; активаторы плазминогена контролируются ингибитором, называемым ингибитором активатора плазминогена (ИАП), активность плазмина ингибируется α2-антиплазмином и α2-макроглобулином, а фибринолиз дополнительно ингибируется активируемым тромбином ингибитора фибринолиза
1. Тромбоциты. Тромбоциты образуются в костном мозге из мегакариоцитов. Мегакариоцитарные клетки-предшественницы (мегакариоцитарные колониеобразующие единицы) делятся, образуя мегакариобласты, которые подвергаются процессу, называемому эндоредупликацией («эндомитотической редупликацией»): при этом происходит деление ядра, но не клетки. Так возникают зрелые мегакариоциты — крупные клетки с несколькими ядрами и цитоплазмой, содержащей тромбоцитарные гранулы. Затем от каждого мегакариоцита отщепляется большое количество тромбоцитов и попадает в кровоток. Образование и созревание мегакариоцитов стимулируются тромбопоэтином, который синтезируется в печени. Тромбоциты циркулируют 8—10 дней, а затем разрушаются в ретикулоэндотелиальной системе. Около 30% периферических тромбоцитов обычно находятся в селезенке и не циркулируют.
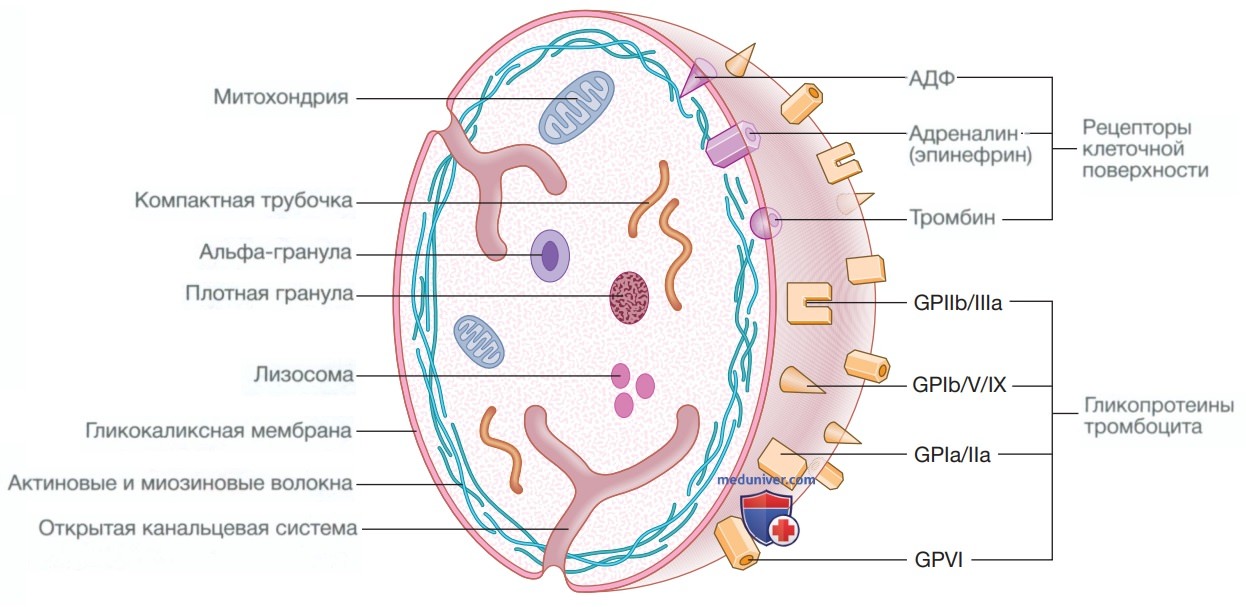
В норме тромбоциты имеют дисковидную форму диаметром 2—4 мкм (рис. 7). Поверхностная мембрана инвагинирует, образуя трубчатую сеть, канальцевую систему, которая обеспечивает канал для эвакуации содержимого гранул после активации тромбоцитов. Препараты, которые ингибируют функцию тромбоцитов и тромбоз, включают ацетилсалициловую кислоту (Аспирин) (ингибитор циклооксигеназы), клопидогрел, прасугрел и тикагрелор (ингибиторы активации, опосредованной аденозиндифосфатом), дипиридамол (ингибитор фосфодиэстеразы) и ингибиторы гликопротеиновых IIb/IIIa рецепторов, абциксимаб, тирофибан и эптифибатид (который предотвращает связывание фибриногена).
Рисунок 7. Структура тромбоцита в норме. Поверхность тромбоцита покрыта гликопротеинами, которые связываются с основными структурами, включая фибриноген, коллаген и фактор Виллебранда, а также рецепторами клеточной поверхности для тромбина, аденозиндифосфата и адреналина (эпинефрина). Через внутренние пути передачи сигналов активация тромбоцита вызывает дегрануляцию альфа-гранул и плотных гранул, что в конечном итоге приводит к агрегации тромбоцитов. Блокада этих путей такими препаратами, как ацетилсалициловая кислота (Аспирин), клопидогрел, тикагрелор, тирофибан и абциксимаб, составляет основу антиагрегантной терапии. ГП — гликопротеин
2. Факторы свертывания крови. Система свертывания крови состоит из каскада растворимых неактивных белковых проферментов, обозначаемых римскими цифрами. После протеолитического расщепления и активации каждый из них способен запускать один или несколько компонентов каскада. Активированные факторы обозначают суффиксом «а». Некоторые из этих реакций требуют присутствия фосфолипидов и кальция. Коагуляция происходит двумя путями: она инициируется внешним (или тканевым фактором) путем и усиливается внутренним путем (см. рис. 6).
Факторы свертывания крови синтезируются в печени, хотя фактор V также вырабатывается тромбоцитами и эндотелиальными клетками. Факторы II, VII, IX и X требуют посттрансляционного карбоксилирования, чтобы они могли участвовать в коагуляции. Фермент карбоксилаза, отвечающий за это в печени, является витамин К-зависимым. В этой реакции витамин К превращается в эпоксид и должен восстановиться до своей активной формы с помощью фермента редуктазы. Данная редуктаза ингибируется варфарином, и это является основой антикоагулянтного эффекта кумаринов. Врожденные (например, гемофилия) и приобретенные (например, печеночная недостаточность) причины дефицита факторов свертывания крови сопровождаются кровотечениями.
Видео №1: лекция кровь как ткань, кроветворение, лимфа
Видео №2: физиология крови (свойства, показатели) - профессор, д.м.н. П.Е. Умрюхин
Видео №3: физиология защитной функции крови (иммунитета) и группы крови - профессор, д.м.н. П.Е. Умрюхин