У пациентов с аллергией или атопией наблюдается изменение реактивности на обычные АГн окружающей среды и пищевые АГн, которые не вызывают клинических реакций у ЗЛ. При клинически выраженной аллергии в организме обычно вырабатываются АТл класса IgE к АГн, вызывающим заболевание. Термин «аллергия» представляет собой клиническое проявление IgE-опосредованных аллергических заболеваний, которые имеют семейную предрасположенность и проявляются гиперреактивностью в органах-мишенях, таких как легкие, кожа, ЖКТ и нос.
Значительное увеличение распространенности аллергических заболеваний за последние несколько десятилетий объясняется изменениями факторов окружающей среды, таких как воздействие табачного дыма, загрязнение воздуха, наличие аллергенов в помещении и на улице, воздействие респираторных вирусов, ожирения и, возможно, снижение частоты некоторых инфекционных заболеваний (гигиеническая гипотеза).
а) Основные элементы аллергических заболеваний:
1. Аллергены. Аллергены почти всегда представляют собой белки, но не все белки являются аллергенами. Чтобы белковый АГн проявлял аллергическую активность, он должен индуцировать продукцию IgE, а это, в свою очередь, должно приводить к развитию реакции ГНТ при последующем контакте с тем же белком. Факторами, которые могут вызвать превращение АГн в аллерген, являются биохим. св-ва аллергена; стимулирующие факторы врожденного иммунного ответа на в-ва аллергены в момент контакта с ними; стабильность аллергена в тканях, ЖКТ, коже или слизистой оболочке; а также доза и продолжительность его пребывания в лимфатических органах во время взаимодействия с иммунной системой.
Это состояние отличается от нормального иммунного ответа на АГн, при котором возникает состояние иммунной реактивности без сопутствующей продукции IgE.
Большинство аллергенов — это белки с молекулярной массой 10-70 кДа. Молекулы с массой <10 кДа не связывают соседние молекулы IgE на поверхности тучных клеток или базофилов. Большинство молекул с массой >70 кДа не проникают через поверхность слизистых, что необходимо для взаимодействия с АГн-презентирующими клетками и стимуляции иммунной системы. Аллергены часто содержат протеазы, которые способствуют дисфункции эпителиального барьера кожи и слизистых оболочек и увеличивают степень проникновения аллергена в ткани организма.
В-ва с низкой молекулярной массой, напр. ЛС, могут становиться аллергенами, реагируя с белками сыворотки крови или белками клеточных мембран. В этом случае иммунная система может их распознать. Углеводородные структуры также м.б. аллергенами и приобретают большое значение с увеличением использования биологических ЛП в клинической практике; пациенты с цетуксимаб-индуцированной анафилаксией имеют специфичные IgE-АТл к галактозе-α-1,3-галактозе.
2. Т-клетки. Все люди контактируют с потенциальными аллергенами. Лица с атопией реагируют на воздействие аллергена быстрой экспансией Т-хелперов (Th; англ. T-helper) 2 типа (Th2), которые секретируют такие цитокины, как IL-4, IL-5 и IL-13, способствуя синтезу IgE и развитию эозинофилии. Аллерген-специфические АТл IgE, связанные с атопическим ответом, можно обнаружить с помощью анализа сыворотки крови или «+» реакции ГНТ на экстракты аллергена при кожной аллергической (скарификационной) пробе.
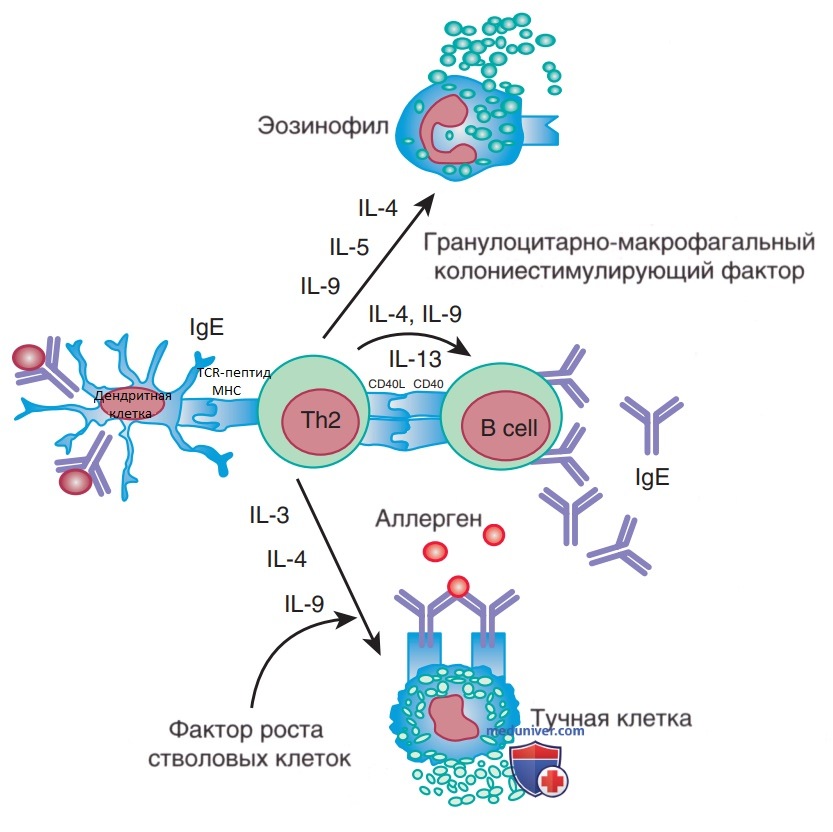
Цитокины Th2 клеток — IL-4 и IL-13 играют ключевую роль в переключении изотипа Ig на IgE-ответ (рис. 1). IL-5 и IL-9 важны для дифференцировки и развития эозинофилов. Комбинация IL-3, IL-4 и IL-9 способствует активации тучных клеток. IL-9 отвечает за выработку слизи. Цитокины Th2-клеток являются важными эффекторными молекулами в патогенезе БА и аллергических заболеваний. Острые аллергические реакции характеризуются инфильтрацией Th2-клеток в пораженных тканях.
Рисунок 1. Роль цитокинов Т-хелперов 2 типа (Th2) в аллергическом каскаде
Кроме того, IL-25, IL-33 и тимический стромальный лимфопоэтин, секретируемые эпителиальными клетками при воздействии аллергенов и респираторных вирусов, вносят определенный вклад со стороны Th2-ответа и эозинофилии.
Часть иммунного ответа на аллерген приводит к пролиферации Th 1 типа (Th1). Клетки Тh1 обычно участвуют в уничтожении в/клеточных микроорганизмов, таких как микобактерии, благодаря способности цитокинов Th1 активировать фагоциты и способствовать выработке опсонизирующих и связывающих комплемент АТл. Компонент ТЫ аллерген-специфического иммунного ответа способствует хронизации и эффекторной фазе аллергического заболевания.
Важным этапом патогенеза в образовании экзематозных высыпаний при атопическом дерматите и слущивании бронхиальных эпителиальных клеток при БА является активация и апоптоз эпителиальных клеток, индуцированных секретируемым Th1-клетками IFN-γ, ФНО-а и лигандом Fas.
Хронические элементы сыпи при аллергических реакциях характеризуются инфильтрацией Th1 и Th17. Это важно, потому что цитокины Th1-клеток, напр. IFN-γ, могут усиливать функцию эффекторных клеток аллергического воспаления (эозинофилов) и тем самым способствовать усилению тяжести заболевания. Клетки Th17 и Th22 связывают иммунный ответ с воспалением ткани; IL-17A, IL-17F и IL-22 являются соответствующими цитокинами-прототипами.
Хотя оба подмножества Т-хелперов играют определенную роль в иммунной защите от внеклеточных бактерий, но IL-17 усиливает воспаление, в то время как IL-22 имеет тканепротективную функцию.
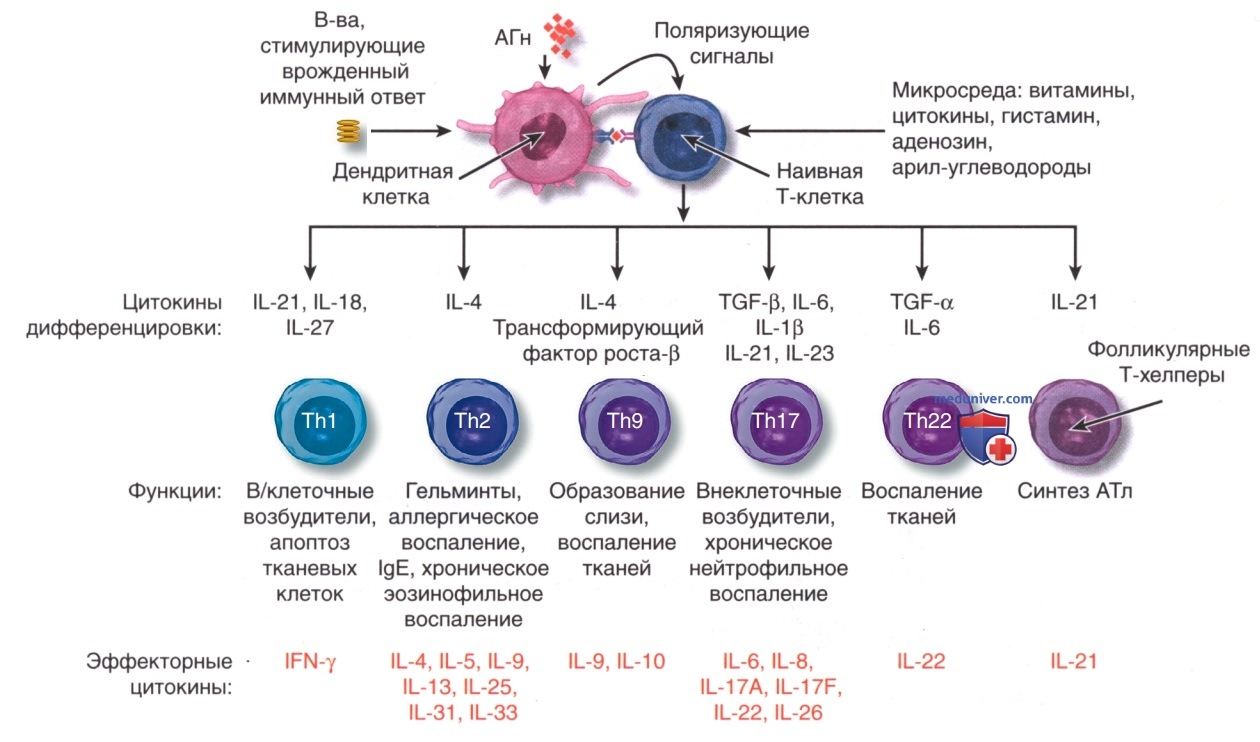
Цитокины семейства IL-17 действуют на несколько типов клеток, включая эпителиальные клетки и АГн-презентирующие клетки, вызывая высвобождение хемокинов, антимикробных пептидов и провоспалительных цитокинов, способствующих усилению воспаления и противомикробных реакций. Кроме того, клетки Th9 продуцируют IL-9, но не др. типичные цитокины Th1, Th2 и Th17, и составляют отдельную популяцию эффекторных Т-клеток, способствующих развитию воспаления тканей. На рис. 2 показаны сложные каскады цитокинов с участием клеток Th1, Th2, Th9, Th17 и Th22.
Рисунок 2. Подмножества эффекторных Т-клеток. После презентации антигена (АГн) дендритными клетками наивные Т-клетки дифференцируются в эффекторные подгруппы Т-хелперов 1 типа (Th1), Т-хелперов 2 типа (Th2), Т-хелперов 9 типа (Th9), Т-хелперов 17 типа (Th17), Т-хелперов 22 типа (Th22) и фолликулярных хелперов. Их дифференцировка требует наличия цитокинов и др. кофакторов, которые высвобождаются из дендритных клеток и также экспрессируются в микросреде. Активация Т-клеток в присутствии интерлейкина IL-4 усиливает дифференцировку и клональную экспансию клеток Th2, поддерживая аллергическую реакцию
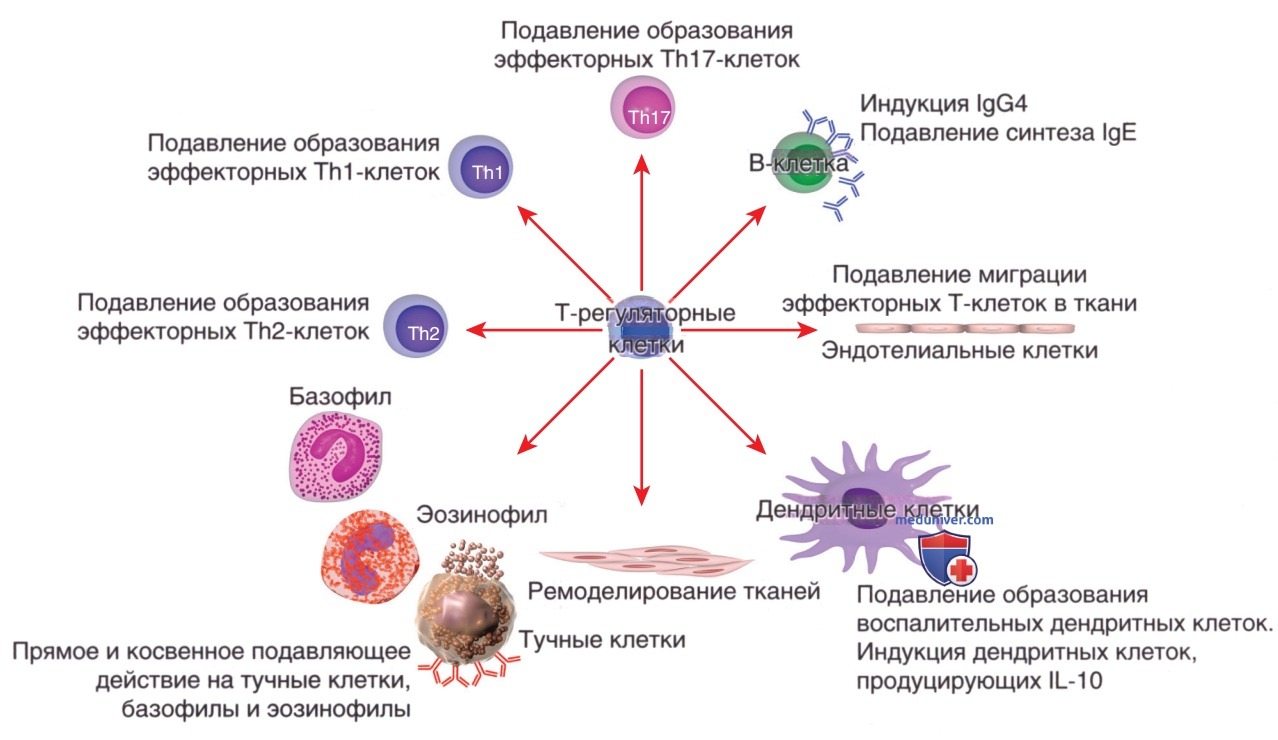
Регуляторные лимфоциты Treg — это подмножество Т-клеток, которые, как считается, играют решающую роль в клинической картине аллергических и аутоиммунных заболеваний. Эти клетки обладают способностью подавлять эффекторные Т-клетки фенотипов Th1, Th2, Th9, Th17 и Th22 (рис. 3). Treg экспрессируют поверхностные молекулы CD4+CD25+ и иммуносупрессивные цитокины, такие как IL-10 и трансформирующий фактор роста-β. Ген фактора транскрипции семейства FOXP3 специфически экспрессируется CD4+CD25+Treg и программирует их развитие и функцию.
Рисунок 3. Контроль аллерген-специфических иммунных ответов. Клетки FoxP3+, CD4+, CD25+ и Tr1 вносят вклад в контроль аллерген-специфических иммунных ответов несколькими основными способами: подавление функции дендритных клеток, которые поддерживают образование эффекторных Т-клеток; ингибирование образования клеток Т-хелперов 1 типа (Th1), Т-хелперов 2 типа (Th2) и Т-хелперов 17 типа (Th17); подавление синтеза аллерген-специфических иммуноглобулинов Е (IgE) и индукция иммуноглобулина G4 (IgG4) и/или иммуноглобулина А; подавление функции тучных клеток, базофилов и эозинофилов; взаимодействие с резидентными тканевыми клетками и ремоделирование, а также подавление миграции эффекторных Т-клеток в ткани
Адаптивный перенос Treg ингибирует развитие эозинофилии ДП и защищает от гиперреактивности бронхов в животных моделях БА. Т-клеточная реакция на аллергены у ЗЛ характеризуется широким диапазоном, от отсутствия поддающегося определению ответа до вовлечения активных механизмов периферической толерантности, опосредованных разл. подмножествами Treg. Лица, не страдающие аллергией, даже несмотря на то что они подвергаются воздействию высоких доз аллергенов, напр. пчеловоды и владельцы кошек, имеют аллерген-специфический IgG4-ответ, сопровождаемый выделением Treg-клеток, продуцирующих IL-10.
Считается, что CD4+CD25+Treg играют важную роль в уменьшении аллергического иммунного ответа, и отсутствие таких клеток может предрасполагать к развитию аллергических заболеваний. У пациентов с мутациями в гене FOXP3 отсутствуют CD4+CD25+Treg, и у них развивается тяжелая иммунная дисрегуляция, сопровождаемая полиэндокринопатией, пищевой аллергией и высокими уровнями IgE в сыворотке крови (болезнь Х-сцепленный аутоиммунно-аллергический синдром дисрегуляции/Х-сцепленный синдром иммунной дисрегуляции, полиэндокринопатии и энтеропатии).
В дополнение к Treg-клеткам, секретирующим IL-10, также и аллерген-специфические Breg-клетки пролиферируют во время проведения аллерген-специфической иммунотерапии и могут играть роль в развитии толерантности к аллергену.
3. Врожденные лимфоидные клетки. Иммунные ответы в популяциях лимфоидных клеток, в которых отсутствуют приспособленные Т- и В-клеточные антигенные рецепторы и поверхностные маркеры миелоидных и лимфоидных клонов, таких как Т-, В-клетки и естественные киллеры, сходны с иммунными ответами ТЫ, Th2 и Thl7/Th22. Эти лимфоидные клетки определяются как врожденные лимфоидные клетки ILC1, ILC2 и ILC3, соответственно, на основании их факторов транскрипции и образования цитокинов. ILC1 в основном образуют IFN-γ; ILC2 продуцируют IL-5, IL-9 и IL-13; а ILC3 синтезируют IL-17 и IL-22 без какой-либо необходимости контакта с АГн/аллергеном.
Убедительные данные указывают на то, что ILC играют важную роль в защите от инфекций и в патогенезе воспалительных заболеваний, таких как БА, аллергические и аутоиммунные заболевания. ILC контролируют среду слизистой оболочки с помощью тесного взаимодействия с эпителиальными клетками и др. тканевыми клетками, продукции цитокинов и индукции хемокинов, которые рекрутируют соответствующие популяции клеток, инициируя и способствуя развитию разл. типов иммунного ответа и воспаления тканей.
ILC2, вероятно, участвуют в индукции БА, аллергического ринита, эозинофильного эзофагита и атопического дерматита за счет активации эпителиальными цитокинами [напр., IL-33, IL-25, тимусный стромальный лимфопоэтин (TSLP; англ. Thymic stromal lymphopoietin)] и взаимодействия с др. иммунными клетками.
4. Антиген-презентирующие клетки. Дендритные клетки, клетки Лангерганса, моноциты и макрофаги обладают способностью представлять аллергены Т-клеткам и тем самым модулировать аллергическое воспаление, контролируя тип развития Т-клеток. АГн-презентирующие клетки представляют собой гетерогенную группу клеток, которые обладают св-вом презентации АГ в рамках HLA МНС и обнаруживаются в основном в лимфоидных органах и коже. Дендритные клетки и клетки Лангерганса уникальны благодаря своей способности праймировать наивные Т-клетки и отвечают за первичный иммунный ответ или фазу сенсибилизации при аллергии.
Считается, что моноциты и макрофаги способствуют активации Т-клеток памяти при повторном воздействии аллергена в фазе реализации аллергической реакции.
Дендритные клетки, расположенные в периферических тканях, таких как кожа, собственная пластинка стенки кишечника и легких, являются относительно незрелыми. Эти незрелые дендритные клетки захватывают АГн в тканях, а затем мигрируют в области скопления Т-клеток в локальных ЛУ. Во время миграции дендритные клетки претерпевают фенотипические и функциональные изменения, характеризующиеся повышенной экспрессией HLA I класса, HLA II класса и костимуляторных молекул, способных взаимодействовать с CD28 Т-лимфоцитов. В ЛУ дендритные клетки представляют переработанные АГн покоящимся Т-клеткам, вызывая их пролиферацию и дифференцировку.
Зрелые дендритные клетки обозначаются как миелоидные или плазматические клетки на основании их способности обеспечивать дифференцировку ТЫ или Th2 соответственно. Критическим фактором, необходимым для поляризации до Thl-клеток, является уровень IL-12, продуцируемый миелоидными дендритными клетками. Напротив, плазматические дендритные клетки характеризуются низкими уровнями IL-12. Плазматические дендритные клетки играют особенно важную роль в противовирусном иммунитете, быстро продуцируя большое количество IFN-a, а также помогают В-клеткам в продукции АТл.
Значительный интерес вызывает роль TSLP, который избыточно экспрессируется на поверхности слизистой оболочки и в коже у лиц с атопией. TSLP усиливает дифференцировку Th2, индуцируя экспрессию OX40L на незрелых миелоидных дендритных клетках в отсутствие продукции IL-12.
Особенность атопических реакций заключается в присутствии на поверхности АГн-презентирующих клеток аллерген-специфических IgE. Важно отметить, что образование комплекса из высокоаффинного рецептора к IgE и аллергена (FcεRI)/IgЕ/аллерген на поверхности АГн-презентирующей клетки значительно облегчает захват и презентацию аллергена. Клиническое значение этого феномена подтверждается тем, что для возникновения экзематозных поражений от аэроаллергенов у пациентов с атопическим дерматитом необходимо присутствие FcεRI-позитивных клеток Лангерганса, несущих молекулы IgE.
Роль низкоаффинного рецептора к IgE (FcεRII, CD23) на моноцитах/макрофагах менее ясна, хотя при определенных условиях он, по-видимому, также может способствовать захвату АГн. Перекрестные связи FcεRII, а также FcεRI на моноцитах/макрофагах приводят к высвобождению медиаторов воспаления. Дендритные клетки играют решающую роль в индукции пероральной толерантности; толерантногенные дендритные клетки распределены в слизистой оболочке и представляют АГн посредством механизма, разработанного для выработки Th1/Treg-супрессивного ответа, который уничтожает аллергенспецифические Т-клетки.
5. Иммуноглобулин Е и его рецепторы. Острая аллергическая реакция зависит от IgE и его способности избирательно связываться с α-цепыо высокоаффинного FcεRI или низкоаффинного FcεRII (CD23). Взаимодействие этих комплексов с аллергеном запускает сложный в/клеточный каскад передачи сигналов, за которым следует высвобождение разл. медиаторов аллергического воспаления из тучных клеток и базофилов. Молекула FcεRI также находится на поверхности АГн-презентирующих дендритных клеток (напр., клеток Лангерганса), но отличается от структуры, обнаруженной на тучных клетках/базофилах, тем, что молекула FcεRI на дендритных клетках не имеет β-цепи.
CD23 обнаруживается на В-клетках, эозинофилах, тромбоцитах и дендритных клетках. Перекрестное сшивание и агрегация FcεRI на тучных клетках и базофилах также может приводить к анафилаксии. Считается, что за этот аберрантный аллергический ответ ответственна разл. экспрессия тирозинкиназ, обеспечивающих положительную и отрицательную регуляцию дегрануляции тучных клеток/ базофилов.
Для индукции синтеза IgE необходимы два основных сигнала. Первый сигнал исходит от IL-4 или IL-13, которые активируют транскрипцию иммуноглобулинового локуса ε в клетках зародышевой линии, тем самым определяя специфичность изотипа. Второй сигнал сводится к взаимодействию CD40 на В-клетках с лигандом CD40 на Т-лимфоцитах. Это активирует механизм рекомбинации, приводящий к переключению синтеза генов ДНК. Взаимодействия между несколькими парами костимулирующих молекул (CD28 и В7; функционально-ассоциированный АГн лимфоцитов-1 и молекула межклеточной адгезии-1; CD2 и CD58) способны дополнительно усиливать первый и второй сигналы с повышением синтеза IgE.
К факторам, угнетающим синтез IgE, относятся цитокины Thl-клеток (IL-12, IFN-α, IFN-γ), IL-10 из Treg, клеток Breg, а также регуляторные дендритные клетки и микробная ДНК, содержащая цитидинфосфат-гуанидиновые повторы.
6. Эозинофилы. Аллергические заболевания характеризуются эозинофилией периферической крови и тканей. Эозинофилы участвуют как во врожденных, так и в адаптивных иммунных ответах и, подобно тучным клеткам, содержат плотные в/клеточные гранулы, которые являются источниками воспалительных белков (см. рис. 1). Эти гранулярные белки включают главный основной белок, эозинофильный нейротоксин, пероксидазу и катионный белок. Белки гранул эозинофилов повреждают эпителиальные клетки, вызывают гиперреактивность ДП и дегрануляцию базофилов и тучных клеток. Главный основной белок, высвобождающийся из эозинофилов, может связываться с кислым фрагментом мускаринового рецептора М2 и блокировать его функцию, тем самым приводя к повышению уровня ацетилхолина и развитию повышенной гиперреактивности ДП.
Эозинофилы также являются источником большого количества Pg и лейкотриенов; в частности, цистеинил-лейкотриен С4 вызывает сокращение гладких мышц ДП и увеличивает проницаемость сосудов. Др. секреторные продукты эозинофилов включают цитокины (IL-4, IL-5, ФНО-α), протеолитические ферменты и активные промежуточные соединения кислорода; все эти в-ва значительно усиливают аллергическое воспаление в тканях.
Некоторые цитокины регулируют функцию эозинофилов при аллергическом заболевании. Эозинофилы развиваются и созревают в костном мозге из миелоидных клеток-предшественников, чувствительных к IL-3, IL-5 и гранулоцитарно-макрофагальныму колониестимулирующему фактору. Контакт с аллергенами у пациентов с аллергией приводит к экспрессии рецептора IL-5 на CD34-клетках костного мозга. Активация рецептора IL-5 вызывает созревание эозинофилов, заставляя эозинофилы синтезировать гранулярные белки, продлевая их выживаемость, усиливая дегрануляцию эозинофилов и стимулируя высвобождение эозинофилов из костного мозга.
Гранулоцитарно-макрофагальный колониестимулирующий фактор также увеличивает пролиферацию и выживаемость клеток, продукцию цитокинов и дегрануляцию эозинофилов. Некоторые хемокины, такие как белки семейства RANTES (регуляция при активации, экспрессия и секреция нормальных Т-клеток), макрофагальный воспалительный белок-1α и эотаксины, играют важную роль в привлечении эозинофилов в локальные аллергические воспалительные реакции тканей. Эотаксины мобилизуют IL-5-зависимые эозинофильные колониеобразующие клетки-предшественники из костного мозга. Эти предшественники быстро выводятся из крови и либо возвращаются в костный мозг, либо попадают в воспаленные участки ткани.
7. Тучные клетки. Тучные клетки образуются из гемопоэтических клеток-предшественников CD34 в костном мозге. После попадания в кровоток они переносятся в периферические ткани, где проходит тканеспецифическое созревание. Развитие и выживание тучных клеток зависит от взаимодействий между рецепторной тирозинкиназой c-kit, экспрессируемой на поверхности тучных клеток, и лигандом фибробластов c-kit, фактором роста стволовых клеток. В отличие от зрелых базофилов, зрелые тучные клетки обычно не циркулируют в кровотоке, а распределены в соединительных тканях, где часто находятся рядом с кровеносными сосудами и под эпителиальными поверхностями, которые контактируют с внешней средой (ДП, ЖКТ и кожа).
Т.о., уже само расположение тучных клеток способствует их участию в аллергических реакциях.
Выделяют, по крайней мере, 2 субпопуляции тучных клеток человека: тучные клетки с триптазой и тучные клетки с триптазой и химазой. Первые располагаются в слизистой оболочке ДП и тонкого кишечника, тогда как вторые преобладают в коже, подслизистом слое ЖКТ и кровеносных сосудах.
Тучные клетки содержат или продуцируют (при соответствующей стимуляции) разнообразные медиаторы, которые по-разному влияют на аллергическое воспаление и функцию органов. К этим в-вам относятся уже имеющиеся в гранулах медиаторы (гистамин, сериновые протеазы, протеогликаны), а также синтезируемые мембранные липиды, цитокины и хемокины. Наиболее важные медиаторы липидной природы — это продукты циклооксигеназного и липооксигеназного путей метаболизма арахидоновой кислоты, которые обладают сильной воспалительной активностью.
Основным продуктом циклооксигеназы тучных клеток является PgD2, а основными продуктами липоксигеназы — сульфидопептидные лейкотриены: LTC4 и его пептидолитические производные LTD4 и LTE4. Тучные клетки также могут продуцировать цитокины, которые способствуют развитию ответа со стороны Th2 (IL-4, IL-13, гранулоцитарно-макрофагальный колониестимулирующий фактор) воспалительной реакции (ФНО-α, IL-6) и регулируют ремоделирование ткани (трансформирующий фактор роста, VEGF). Иммунная активация тучных клеток и базофилов обычно начинается с перекрестного сшивания IgE, связанного с FcεRI, с поливалентным аллергеном. Содержание FcεRI на поверхности тучных клеток увеличивается за счет действия IL-4 и IgE.
С терапевтической точки зрения представляет собой интерес, что при введении АТл против IgE, снижающих его уровень в сыворотке крови, количество FcεRI на поверхности тучных клеток уменьшается.
б) Механизмы аллергического воспаления тканей. IgE-опосредованные иммунные ответы можно с хронологической точки зрения классифицировать на 3 типа реакций. Немедленная реакция на попадание аллергена в органы-мишени, обусловленная дегрануляцией тучных клеток и высвобождением готовых медиаторов воспаления. Она развивается за 1-30 мин после контакта с аллергеном и разрешается за 1-3 ч. Немедленные реакции связаны с локальным повышением проницаемости сосудов, что приводит к утечке белков плазмы, отеку тканей и усилению кровотока, а также к зуду, чиханию, бронхообструкции и острым спастическим болям в животе, в зависимости от пораженного органа.
Вторая, поздняя реакция может развиваться через несколько часов после воздействия аллергена, достигая максимальной выраженности через 6-12 ч и разрешается через 24 ч. Поздняя фаза в коже характеризуется отеком, покраснением и уплотнением; в носу — длительной заложенностью носа; в легких — ОДП и стойкими сухими хрипами. Как правило, поздняя фаза ответа связана с ранней инфильтрацией нейтрофилами и эозинофилами, вслед за которыми активируются базофилы, моноциты, макрофаги и 1112-лимфоциты. Для этого необходима усиленная экспрессия молекул адгезии на поверхности воспалительных клеток, а также лигандов этих молекул на эндотелиальных клетках, которая регулируется цитокинами.
Через несколько часов после воздействия аллергена ФНО-α, высвобождаемый активированными тучными клетками, индуцирует экспрессию молекул клеточной адгезии на эндотелии, что облегчает проникновение разл. воспалительных клеток через эндотелий.
Преимущественное накопление эозинофилов происходит за счет взаимодействий между молекулами селективной адгезии на поверхности эозинофильных клеток (напр., интегрином α4β1 или очень поздним АГн-4), экспрессия которой на эндотелиальных клетках усиливается под действием IL-4 и IL-13. ILC2 получают такие сигналы от эпителиальных клеток, как IL-33, TSLP и IL-25, активируются и начинают высвобождать свои цитокины IL-5 и IL-13, инициируя иммунный ответ 2 типа.
Хемокины — это хемотаксические цитокины, которые играют центральную роль в тканевой миграции воспалительных клеток. Белки RANTES, макрофагальный воспалительный белок (MIP-1α), белки хемотаксиса моноцитов МСР-3 и МСР-4 являются хемоаттрактантами для эозинофилов и мононуклеарных клеток, тогда как эотаксины относительно селективны именно в отношении эозинофилов.
Эти хемоаттрактанты были обнаружены в эпителии, макрофагах, лимфоцитах и эозинофилах в местах реакций поздней фазы и аллергического воспаления тканей. Блокада действий этих хемокинов приводит к значительному снижению направленной миграции эффекторных аллергических клеток в ткани.
При реакции 3 типа, хронических аллергических заболеваниях, воспаление тканей может сохраняться от нескольких дней до нескольких лет. Ряд факторов способствуют стойкому воспалению тканей, включая повторяющийся контакт с аллергенами и микроорганизмами. Повторная стимуляция аллергических эффекторных клеток, таких как тучные клетки, базофилы, эозинофилы и клетки Th2, способствует отсутствию разрешения воспалительного состояния. Кроме того, цитокины Th2-типа (IL-3, IL-5, гранулоцитарно-макрофагальный колониестимулирующий фактор), секретируемые во время аллергических реакций, могут продлевать выживаемость аллергических эффекторных клеток, замедляя апоптоз.
Локальная дифференцировка предшественников эозинофилов, инфильтрирующих ткани, под действием IL-5 приводит к спонтанному образованию эозинофилов, дополнительно поддерживая местное повреждение тканей. Ремоделирование тканей, приводящее к необратимым изменениям в органах-мишенях, также является признаком хронических аллергических заболеваний. При БА ремоделирование включает утолщение стенок ДП и подслизистого слоя, а также гипертрофию и гиперплазию гладких мышц, что сопровождается ухудшением функции легких. Это неожиданная роль эозинофилов в ремоделировании ДП, а также в хроническом воспалении. При атопическом дерматите лихенификация является очевидным признаком ремоделирования кожи.
Как правило, считается, что иммунная реакция 2 типа лежит в основе большинства случаев БА, атопического дерматита, хронического риносинусита и аллергического ринита как общая характеристика иммунного/воспали-тельного ответа. В иммунной реакции 2 типа участвуют клетки Th2, В-клетки 2 типа, ILC2, IL-4, образующие естественные киллеры Т-клетки, базофилы, эозинофилы и тучные клетки, а также их основные цитокины. Из всей сложной сети цитокинов IL-4, IL-5, IL-9 и IL-13 в основном секретируются клетками иммунной системы, a IL-25, IL-31, IL-33 и TSLP — клетками тканей, особенно эпителиальными клетками.
Многие АГн, связанные с БА, такие как протеазные аллергены, грибковые экстракты и вирусные инфекции, стимулируют выработку IL-33, TSLP и IL-25 клетками эпителия и разл. иммунными клетками и вызывают эозинофильное астматическое воспаление ДП за счет активации ILC2 в легких.
С др. стороны, IL-31 играет роль в зуде при атопическом дерматите. Цитокины Th2 не только поддерживают аллергическое воспаление, но также влияют на ремоделирование тканей, активируя резидентные клетки в органах-мишенях; IL-4, IL-9 и IL-13 вызывают гиперсекрецию слизи и метаплазию клеток слизи; IL-4 и IL-13 стимулируют рост фибробластов и синтез белков внеклеточного матрикса; a IL-5 и IL-9 увеличивают субэпителиальный фиброз. Трансформирующий фактор роста-β, синтезируемый эозинофилами и фибробластами, также может усиливать субэпителиальный фиброз.
IL-11, экспрессируемый эозинофилами и эпителиальными клетками, способствует фиброзу подслизистого слоя, усиливая отложение коллагена и накопление фибробластов. Возникающее в результате повреждение тканей усиливает дальнейшее нарушение целостности эпителия за счет высвобождения провоспалительных цитокинов, отложения внеклеточного матрикса в органах-мишенях и ангиогенеза. Генетическая предрасположенность к аберрантным реакциям заживления может способствовать хронизации заболевания. Т.о., однажды возникнув, аллергическая реакция сама себя поддерживает благодаря общей иммунной реакции 2 типа и приводит к хроническому заболеванию у людей с генетической предрасположенностью.
Последующая инфильтрация Th1- и Th17-клетками усиливает воспалительный потенциал аллергических эффекторных клеток и способствует сохранению хронической воспалительной реакции тканей за счет высвобождения провоспалительных цитокинов и хемокинов. Кроме того, аутоиммунный ответ м.б. причиной аллергического воспаления в результате возможных механизмов действия через аутоАТл IgE, аутоАТл IgG и аутореактивность Th1-клеток и Th17-клеток.
в) Генетическая основа атопии. Аллергические заболевания — это сложные генетические состояния, в развитии которых играют роль факторы окружающей среды. Несколько важных групп генов связаны с аллергическими заболеваниями: гены, которые регулируют системную экспрессию атопии (повышенный синтез IgE, эозинофилия, реакции тучных клеток) и обычно экспрессируются при разл. аллергических заболеваниях, гены, которые контролируют барьерную функцию в определенных органах-мишенях (напр., кожа при атопическом дерматите, легкие при БА, ЖКТ при пищевой аллергии) и гены, кодирующие рецепторы распознавания объектов врожденной иммунной системы, которые взаимодействуют с микроорганизмами и влияют на адаптивные иммунные ответы. После инициирования аллергических реакций генетическая предрасположенность к хроническому аллергическому воспалению и аберрантный процесс заживления способствуют ремоделированию тканей и персистированию заболевания.
Атопические заболевания характеризуются выраженной семейной предрасположенностью с 60% наследуемостью в близнецовых исследованиях БА и атопического дерматита. Область 5q23-35 включает несколько генов, участвующих в патогенезе аллергических заболеваний, в т.ч. гены, кодирующие цитокины Th2 (IL-3, IL-4, IL-5, IL-9, IL-13, гранулоцитарно-макрофагальный колониестимулирующий фактор). Среди них ген IL4 — хорошо изученный потенциальный ген-кандидат. Замена нуклеотида в положении 589 промоторной области IL4 приводит к образованию уникального участка связывания для фактора транскрипции нуклеарный фактор-АТл, повышению транскрипции гена IL-4, более высокой аффинности связывания нуклеарного фактора-АТл и увеличению образования IgE.
Точно так же варианты кодирующей области IL13 были связаны с БА и атопическим дерматитом. Обнаружена также связь между атопией и усиливающим функцию полиморфизмом гена а-субъединицы рецептора IL-4, который расположен на 16 хромосоме. Это открытие согласуется с важной ролью IL-4, IL-13 и их рецепторов в иммунопатогенезе аллергических заболеваний.
В полногеномных исследованиях также обнаружена ассоциация атопии с областью хромосомы 11q13. Ген, кодирующий β-субъединицу FcεRI-β, был определен как предположительный ген-кандидат в этой области. Ген β-субъединицы модифицирует активность FceRI на тучных клетках, и несколько генетических вариантов FcεRI-β связаны с БА и атопическим дерматитом. Хромосома 6 содержит гены, кодирующие HLA I класса и II класса, которые регулируют специфичность и интенсивность иммунных ответов на определенные аллергены. IgE-ответы на специфические аллергены, такие как АГн амброзии Amb а V и аллерген пылевого клеща Der р I, были связаны со специфическими локусами HLA II класса.
На 6-й хромосоме также находится ген ФНО-α, ключевого цитокина, который способствует миграции воспалительных клеток. Полиморфизм гена ФНО-α связан с БА. Недавнее полногеномное исследование показало, что генетические полиморфизмы в гене, кодирующем IL-33, который является основным активатором ILC2, и его рецепторе IL-1RL1 (ST2) тесно связаны с развитием БА.
Дисфункция защитного барьера играет ключевую роль в патогенезе аллергических заболеваний. Исследования генетических взаимосвязей атопического дерматита продемонстрировали важность хромосомы 1q21, которая содержит кластер генов, участвующих в дифференцировке эпидермиса. Филаггрин — это белок, который имеет важное значение в формировании рогового слоя. Нуль-мутации гена филаггрина тесно связаны с ранним началом и тяжелым течением атопического дерматита. Было показано, что мутации в гене, кодирующем ингибитор сериновой протеазы SPINK5, вызывают болезнь Нетертона, моногенное расстройство, связанное с эритродермией, пищевой аллергией и высоким сывороточным уровнем IgE. Частый полиморфизм SPINK5 (в частности, Glu420Lys) увеличивает риск развития атопического дерматита и БА.
SPINK5 экспрессируется в наружном слое эпидермиса и считается критически важным для нейтрализации протеолитической активности Staphylococcus aureus и распространенных аллергенов, таких как Der р I, которые используют эти протеазы для проникновения через кожу, вызывая аллергические реакции.
Дисфункция барьера важна и при др. аллергических заболеваниях, таких как БА и риносинусит, но, вероятно, при этом изменяются др. барьерные гены, например, те, которые кодируют щелевые контакты.
Гены-кандидаты, связанные с повышением предрасположенности к БА, были идентифицированы с помощью позиционного клонирования: GPRA (рецептор, связанный с G-белком, с увеличением предрасположенности к БА на хромосоме 7р14), ADAM-33 (дезинтегрин и металлопротеиназа 33 на хромосоме 20р) и DPP10 (дипептидилпептидаза 10 на хромосоме 2q14). Функции этих генов не соответствуют классическим путям атопии и, следовательно, позволяют по-новому взглянуть на патогенез БА.
GPRA кодирует рецептор, связанный с G-белком, изоформы которого экспрессируются в эпителиальных клетках бронхов и гладких мышцах у пациентов с БА, что указывает на важную роль этих тканей при данном заболевании. ADAM-33 экспрессируется в гладких мышцах бронхов и связан с гиперреактивностью бронхов. DPP10 кодирует дипептидилдипептидазу, которая может удалять 2 концевых пептида из некоторых провоспалительных хемокинов; это изменение способно модулировать аллергическое воспаление.
Рецепторы распознавания образов врожденной иммунной системы, которые экспрессируются эпителиальными клетками и дендритными клетками, связаны с повышенной предрасположенностью к заболеванию. Эти рецепторы распознают определенные микробные компоненты. Полиморфизм CD14 (связывает эндотоксин), Toll-подобного рецептора 2 типа (который связывает Staphylococcus aureus), а также Т-клеточного иммуноглобулинового домена и муцинового домена (который связывает HAV) коррелирует с повышением предрасположенности к БА или/и атопическому дерматиту. Нарушение регуляции этих первых систем иммунной защиты может привести к патологической реакции на распространенные аллергены окружающей среды.