Избыточная экспрессия мерлина, в отличие от других представителей белков семейства ERM, вызывает подавление роста. В дополнение к актину цитоскелета, мерлин продемонстрировал связь с клеточной мембраной доменов, которые являются многостимульными сигнальными молекулами, регулирующими клеточный ответ на пролиферативные и антипролиферативные стимулы.
Клетки вестибулярной шванномы с инактивацией НФ2 имеют резкие изменения в клеточном распределении. В настоящее время определены некоторые белки, способные взаимодействовать с мерлином, к ним относятся: семейство ERM белков, CD44, F-актин, паксиллин, микротрубочки, βII-спектрин, β1-интегрин, β-фодрин, кофактор регуляции Na+-H+ обменника (NHE-RF), SCHIP-1, регулируемые фактором роста гепатоцитов субстраты тирозинкиназы (HRS), р21-активатор киназы 1 и 2 (Pakl и Pak2), Rac1, субъединица RIB протеинкиназы А, компоненты кадгерин-опосредованной клеточной адгезии, PIKE-L [усилитель фосфатидилинозитол-3-киназы (РI3-киназа)] и эрбин (erbB2-взаимодействующий белок).
В настоящее время нет четкого объяснения, как все эти межбелковые взаимодействия обеспечивают супрессию опухоли мерлином. Соединение мерлина с CD44 и β1-интегрином повышает вероятность функционирования мерлина как молекулярного переключателя в сигнальных путях. CD44 является рецептором трансмембранной гиалуроновой кислоты, который участвует в межклеточной адгезии, клеточно-матриксной адгезии, подвижности клеток и метастазировании.
Мерлин опосредует контактное ингибирование клеточного роста сигнализированием из внеклеточного матрикса. При высокой плотности клеток мерлин гипофосфорилируется и подавляет клеточный рост в ответ на окружающие клетки мукополисахариды. При низкой плотности клеток мерлин фосфорилируется, способствует росту, и образует комплексы с эзрином, моэзином и CD44. Эти данные указывают на то, что мерлин и CD44 образуют молекулярный переключатель, который определяет приостановку роста клеток или их пролиферацию.
Исследования Lallemand и соавт. показали, что в фибробластах эмбриона мыши Нф2-/-, дефицит Нф2 привел к скоплению клеток, гиперпролиферации и дефектным кадгерин-опосредованным межклеточным взаимодействиям. При нормальном функционировании эти взаимодействия прекращают пролиферацию при остановке деления клеток после контактов в специфических соединениях, называемых «прилипающими». Кроме того, колоконизация и взаимодействие мерлина вызывает созревание этих адгезивных соединительных компонентов с соприкосновением или соседством клеток дикого типа.
Иначе говоря, при нормальном взаимодействии шванновских клеток друг с другом специфическое сигнализирование через эти соединения останавливает дальнейший рост. В отсутствие мерлина адгезивные соединения являются незрелыми и не приостанавливают завершение проста шванновских клеток, когда они контактируют. Эти результаты указывают на то, что функции мерлина в качестве опухолевого супрессора частично контролируются кадгерин-опосредованным межклеточным взаимодействием.
Функционирование мерлина регулируется фосфорилированием. Было продемонстрировано, что Rac1, член семейства Rho ГТФазы, содействует фосфорилированию мерлина, тем самым, инактивирует его механизм подавления роста. Показано взаимодействие мерлинома с эффекторами Rac1 и Cdc42 ГТФазы, членами семейства р21-активированный киназы (РАК). В частности, (Рак2) продемонстрировал фосфорилирование мерлина в позиции серина 518 и инактивацию его функции.
Kissil и соавт. сообщают также о взаимодействии между мерлином и Paid и способности мерлина препятствовать активизации Pak1. Потеря экспрессии мерлина приводит к неадекватной активации Pak1, в то время как его избыточная экспрессия — к ингибированию деятельности Pak1. Напротив, мерлин активируется дефосфорилированием серина в позиции 518, которое происходит в сыворотке или на межклеточных и клеточно-матриксных контактах. Хотя члены семейства РАК участвуют в различных клеточных процессах, таких как реорганизация цитоскелета и передача сигнала апоптоза, их точная роль и функции не определены. Jin и соавт. определили миозин фосфатазу (MYPT-1-PP15) в качестве мерлин фосфатазы. Интересно, что клеточный MYPT- 1-РР15-специфический ингибитор, CPI-17, может привести к снижению активности мерлина путем его фосфорилирования, Ras-активации и трансформации.
Эти результаты предполагают MYPT-1-PP16 и CPI-17 в качестве важных регуляторных компонентов в направлении супрессии опухолевого роста мерлином.
Мерлин как регулятор роста при конформации и межбелковом взаимодействии. Деятельность ERM белков контролируется самоассоциацией N- и С-концевых участков белков. Белки ERM могут существовать в «закрытой» конформации, где N- и С-концевые участки подвергаются внутримолекулярным взаимодействиям, таким образом, формируется белок маскирующий стабилизированный актин-связывающий участок. Молекулы могут быть преобразованы в «открытую» конформацию, в которой внутримолекулярное взаимодействие нарушает сигналы, такие как фосфорилирование или лечение с фосфоинозитидом.
Функционирование мерлина в качестве регулятора роста связано и с его способностью образовывать подобные внутримолекулярные связи. Выявлено два таких взаимодействия. При первом взаимодействии происходит сворачивание остатков внутри N-концевого участка белка, а при втором взаимодействии весь белок сворачивается с соединением N- и С-концевых участков белка. По сходному с белками ERM типу мерлин проходит цикл между «открытой» и «закрытой» конформацией, что и определяет его связь с ERM белками или с другими молекулами, преобразующими тормозной сигнал роста мерлина. Кроме того, возможно, что скручиванием мерлина регулируется связь между мерлином и HRS, субстратом, включенным в сигнальный путь инициацией фактора роста гепатоцитов (HFS), связанных с c-met рецептором.
Эти результаты позволяют предположить, что способность мерлина к циклам от «открытой» к «закрытой» конформации может включать сигнальные пути CD44 и HGF. Все эти данные связаны с регуляцией роста. Кроме того, мерлин может осуществлять свою деятельность за счет ингибирования фосфатидилинозитол-3-киназы (PI3-киназы) посредством связывания с PIKE-L, мозговой специфической ГТФазы, которая связывает PI3-киназу и стимулирует его липидкиназную активность. Этот факт позволяет предположить, что PIKE-L является важным медиатором подавления роста мерлина. В качестве дополнения мы обнаружили, что PI3-киназы/Akt пути активируется в вестибулярной шванноме (ВШ).
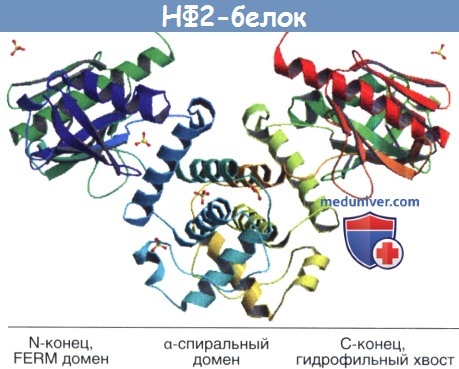
НФ2 белок имеет три крупные части.
За FERM доменом следуют альфа-спиральный домен и гидрофильный хвост.
Мерлин может димеризоваться с собой и гетеродимеризоваться с другими белками семьи ERM.
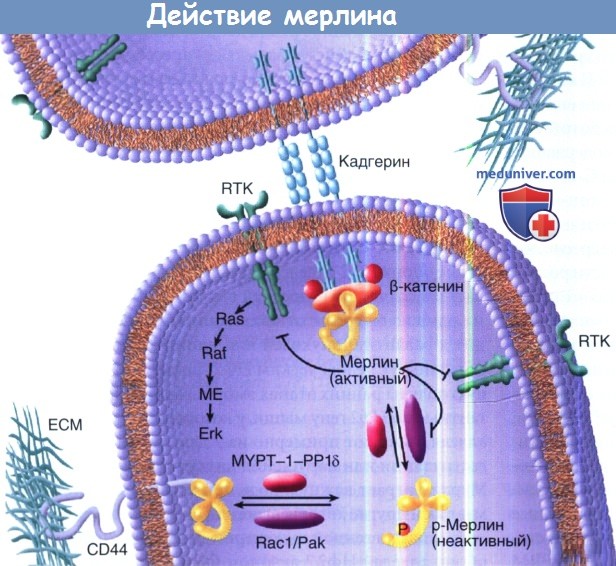
Схематическое изображение действия мерлина.
Схема показывает, как Rac1 и Pak способствуют преобразованию мерлина из закрытой конформации белка в открытую путем его фосфорилирования.
В результате в открытой конформации мерлин может взаимодействовать с CD44 и содействовать связыванию актина цитоскелета в составе клеточной мембраны.
Показана инактивация функции мерлина при его фосфорилировании в 518 позиции серина р21-активатор киназой-2 (Рак2).
Мерлин активируется дефосфорилированием миозин-фосфатазой (MYPT-1-PP16).