Функциональная анатомия и физиология секреции инсулина, контроля уровня глюкозы в крови
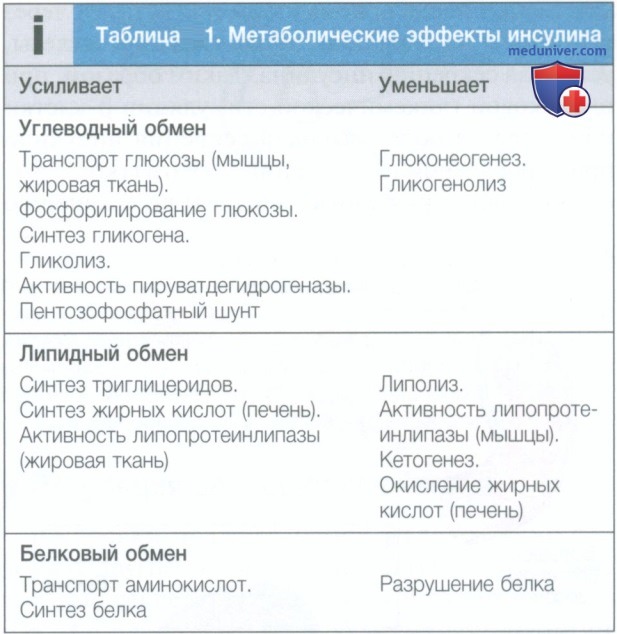
а) Регуляция секреции инсулина. Инсулин представляет собой основной регулятор метаболизма и накопления глюкозы (табл. 1), он секретируется β-клетками поджелудочной железы в систему воротной вены (рис. 1). β-Клетки поджелудочной железы способны точно регулировать концентрацию глюкозы в крови благодаря сопряжению глюкозы и других алиментарных стимулов с секрецией инсулина (см. рис. 1). Поступление глюкозы в β-клетку поджелудочной железы осуществляется путем диффузии по градиенту концентрации с помощью переносчиков глюкозы в клеточной мембране (GLUT).
Рисунок 1. Структура и эндокринная функция поджелудочной железы. А — в норме поджелудочная железа взрослого человека содержит около 1 млн островков, которые рассеяны по всей паренхиме экзокринной части. Гистологическая структура показана на рис. ниже. В — ядро каждого островка состоит из β-клеток, которые вырабатывают инсулин, и окружено корой из эндокринных клеток, синтезирующих другие гормоны, в том числе глюкагон (α-клетки), соматостатин ( δ-клетки) и панкреатический полипептид (РР-клетки). С — схематическое изображение β-клетки поджелудочной железы. 1. Глюкоза поступает в клетку с помощью переносчика глюкозы (GLUT1 или GLUT2). 2. Затем глюкоза подвергается гликолизу, а последующее окислительное фосфорилирование в митохондриях приводит к увеличению внутриклеточного содержания аденозинтрифосфата. 3. Аденозинтрифосфат вызывает закрытие чувствительного к аденозинтрифосфату калиевого канала (который состоит из четырех субъединиц Kir6.2 и четырех субъединиц SUR1). Это приводит к деполяризации мембраны. 4. Повышение мембранного потенциала вызывает входящий ток кальция из-за открытия потенциалозависимых кальциевых каналов. Это повышение внутриклеточного уровня кальция индуцирует слияние секреторных гранул инсулина с клеточной мембраной, что приводит к секреции инсулина. 5. Другие стимулы, такие как глюкагоноподобный пептид-1 или глюкозозависимый инсулинотропный пептид, действуют на рецепторы, связанные с G-белком, увеличивая уровень циклического аденозинмонофосфата и усиливая секрецию инсулина. Генетические дефекты в β-клетке приводят к сахарному диабету. Первично поражаются гены глюкокиназы (начальная стадия гликолиза) и HNF1a, HNF4a и HNF10 (ядерные факторы транскрипции). Две группы лекарственных препаратов воздействуют на β-клетку, стимулируя секрецию инсулина. Производные сульфонилмочевины закрывают чувствительные к аденозинтрифосфату калиевые каналы, вызывая деполяризацию мембраны, входящий ток кальция и секрецию инсулина. Инкретиномиметики увеличивают концентрацию эндогенного глюкагоноподобного пептида 1 и глюкозозависимого инсулинотропного пептида (ингибиторы дипептидилпептидазы 4) или действуют непосредственно на рецептор глюкагоноподобного пептида 1 (агонисты рецепторов глюкагоноподобного пептида 1). Обе эти группы препаратов увеличивают секрецию инсулина, но только после первоначального стимула в форме закрытия чувствительных к аденозинтрифосфату калиевых каналов β-клеток глюкозой (или производными сульфонилмочевины)
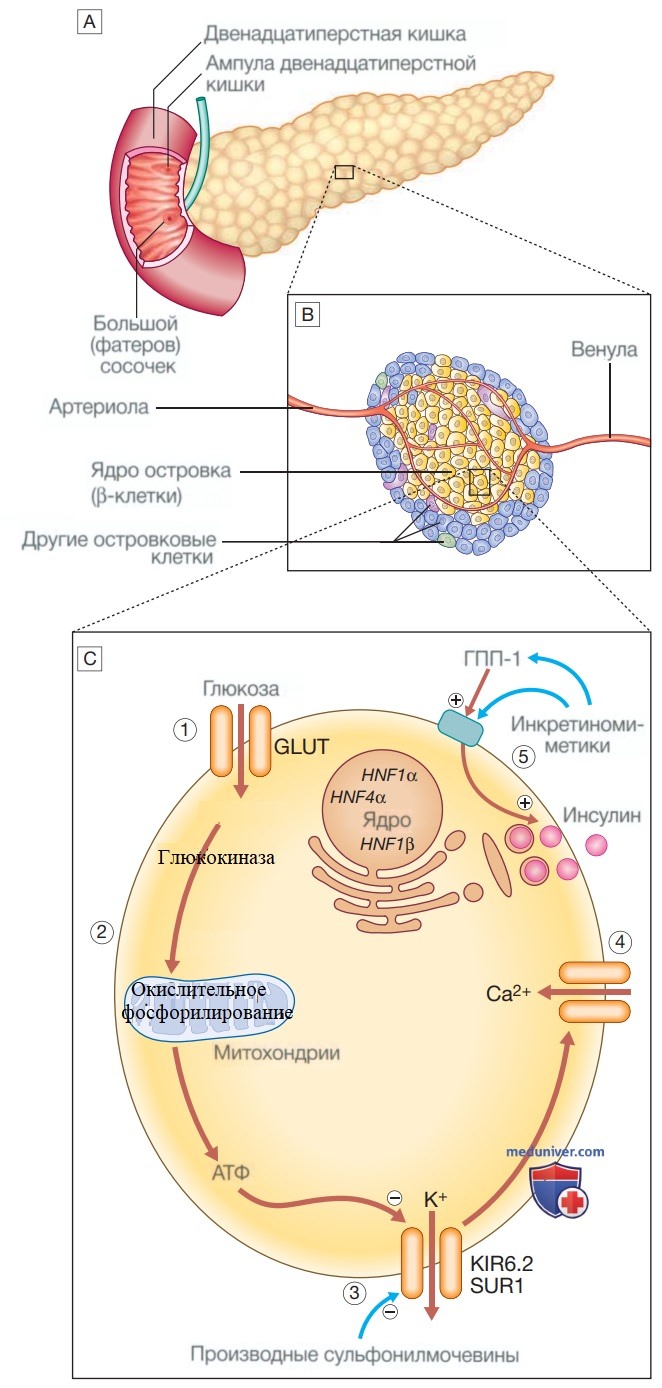
Патогенез сахарного диабета 1-го типа. Предполагаемая последовательность событий при развитии сахарного диабета 1 -го типа. Внешние триггеры описаны в тексте
Затем глюкоза метаболизируется за счет гликолиза и окислительного фосфорилирования. Первая стадия гликолитического пути, превращение глюкозы в глюкозо-6-фосфат, катализируется ферментом глюкокиназой. Глюкокиназа обладает низким сродством к глюкозе, поэтому ее активность в нормальных физиологических условиях заметно различается в зависимости от концентрации глюкозы; это превращает глюкокиназу в очень эффективный сенсор глюкозы в β-клетке. При классическом прямом или запускающем пути метаболизм глюкозы приводит к увеличению внутриклеточного содержания АТФ и снижению уровня аденозиндифосфата, что вызывает закрытие чувствительного к АТФ калиевого канала (КАТФ).
В результате деполяризация мембраны β-клетки приводит к секреции инсулина из-за инициации высвобождения кальция через потенциалозависимые кальциевые каналы. Кроме этого пути, количество высвобождаемого инсулина может увеличиваться под действием фонового уровня глюкозы в крови, других питательных веществ и пептидов, а также нейрогенно — через симпатическую и парасимпатическую нервную систему. Хорошим примером такого усиления высвобождения инсулина является секреция двух кишечных пептидов после приема пищи.
ГПП-1 и глюкозозависимый инсулинотропный пептид высвобождаются из L- и К-клеток желудочно-кишечного тракта соответственно после приема пищи и действуют через рецепторы на β-клетках поджелудочной железы, усиливая секрецию инсулина.
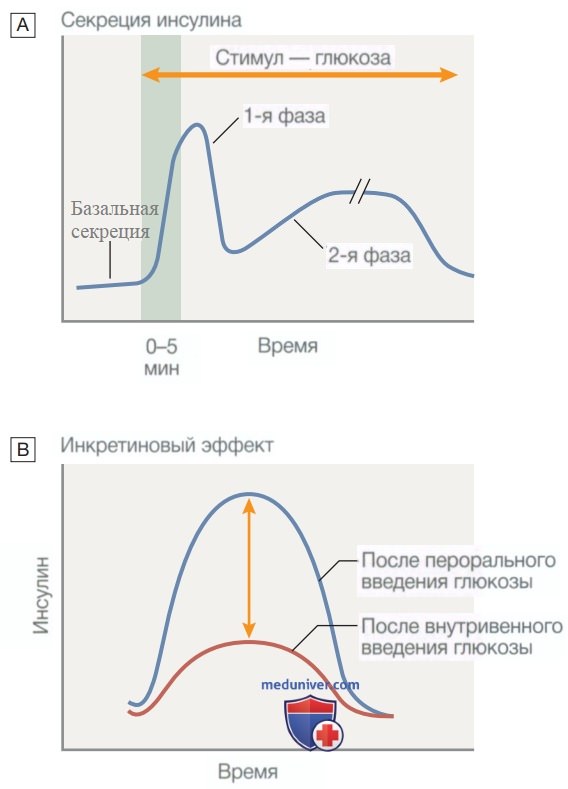
Таким образом, при одинаковой гликемической стимуляции β-клетки наблюдается более высокая секреция инсулина при пероральном поступлении глюкозы (когда высвобождаются кишечные пептиды) по сравнению с внутривенным введением декстрозы (Глюкозы) (которое не стимулирует секрецию кишечных пептидов).
Это усиление секреции инсулина после перорального приема декстрозы (Глюкозы) называется инкретиновым эффектом (рис. 2), а ГПП-1 и глюкозозависимый инсулинотропный пептид называют гормонами из семейства инкретинов.
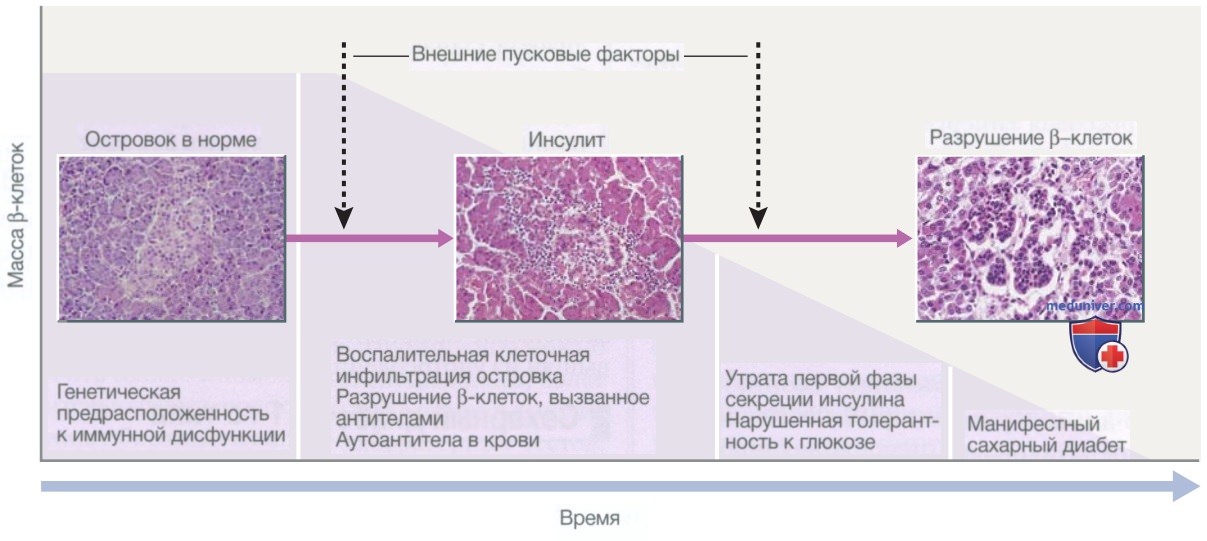
Рисунок 2. Секреция инсулина в ответ на внутривенное или пероральное введение декстрозы (Глюкозы). А — немедленная первая фаза секреции инсулина происходит в ответ на повышение уровня глюкозы в крови, после чего следует длительная вторая фаза. В — инкретиновый эффект объясняет более выраженную секрецию инсулина при пероральном поступлении декстрозы (Глюкозы) по сравнению с внутривенным введением декстрозы (Глюкозы*) для достижения такого же повышения концентрации глюкозы в крови. Дополнительный стимул к секреции инсулина опосредуется высвобождением пептидов из кишечника, и эти эффекты используются при терапии инкретиномиметиками
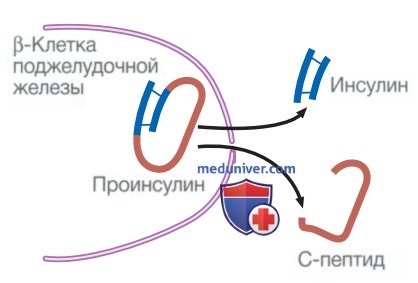
Инсулин синтезируется в виде предшественника гормона (проинсулин), который состоит из α- и β-цепей, соединенных С-пептидом (рис. 3). С-пептид расщепляется β-клеточными пептидазами с образованием инсулина (который теперь состоит из α- и β-цепей) и свободного С-пептида. Секреция инсулина в ответ на поступление глюкозы традиционно происходит в две фазы (см. рис. 2). Первая, быстрая фаза представляет собой секрецию предварительно образованного инсулина из гранул внутри β-клеток, тогда как более продолжительная вторая фаза обусловлена синтезом нового инсулина.
Рисунок 3. Расщепление проинсулина с образованием инсулина и С-пептида. Проинсулин в β-клетке поджелудочной железы расщепляется с высвобождением инсулина и эквимолярных количеств инертного С-пептида (связывающего пептида). Измерение уровня С-пептида можно использовать для оценки секреторной способности β-клеток
б) Регуляция секреции глюкагона. Островки поджелудочной железы также содержат другие эндокринные клетки — α-клетки, которые секретируют пептидный гормон глюкагон, и β-клетки, которые образуют соматостатин (см. рис. 1, В). α-Клетки составляют около 20% популяции островковых клеток у человека. Глюкагон оказывает действие, противоположное инсулину, и стимулирует гликогенолиз в печени (и почках), что приводит к увеличению образования глюкозы в печени. Регуляция секреции глюкагона α-клетками сложна, но полагают, что важную регуляторную роль играют секреция инсулина β-клетками, совместная секреция цинка и γ-аминомасляной кислоты, а также соматостатина из δ-клеток.
Это означает, что инсулин и глюкагон строго и реципрокно регулируются таким образом, что отношение инсулина к глюкагону в воротной вене является основным фактором, определяющим образование глюкозы в печени. Глюкагон также крайне важен для защиты организма от гипогликемии.
в) Гомеостаз глюкозы в крови. Уровень глюкозы в крови строго регулируется и поддерживается в узком диапазоне. Это важно для обеспечения непрерывного поступления глюкозы в ЦНС. Головной мозг способен лишь немного накапливать энергию в форме гликогена и триглицеридов, а гематоэнцефалический барьер в значительной степени является непроницаемым для жирных кислот, поэтому головному мозгу требуется постоянное поступление глюкозы из печени в целях окисления и, следовательно, образования АТФ. Гомеостаз глюкозы достигается благодаря скоординированной работе различных органов, но в основном он обусловлен балансом между поступлением глюкозы в кровоток из печени, дополняемый всасыванием глюкозы в кишечнике после приема пищи, и захватом глюкозы периферическими тканями, особенно скелетными мышцами и головным мозгом.
После приема пищи, содержащей углеводы, нормальный уровень глюкозы в крови поддерживается за счет:
• подавления образования глюкозы в печени;
• стимуляции захвата глюкозы печенью;
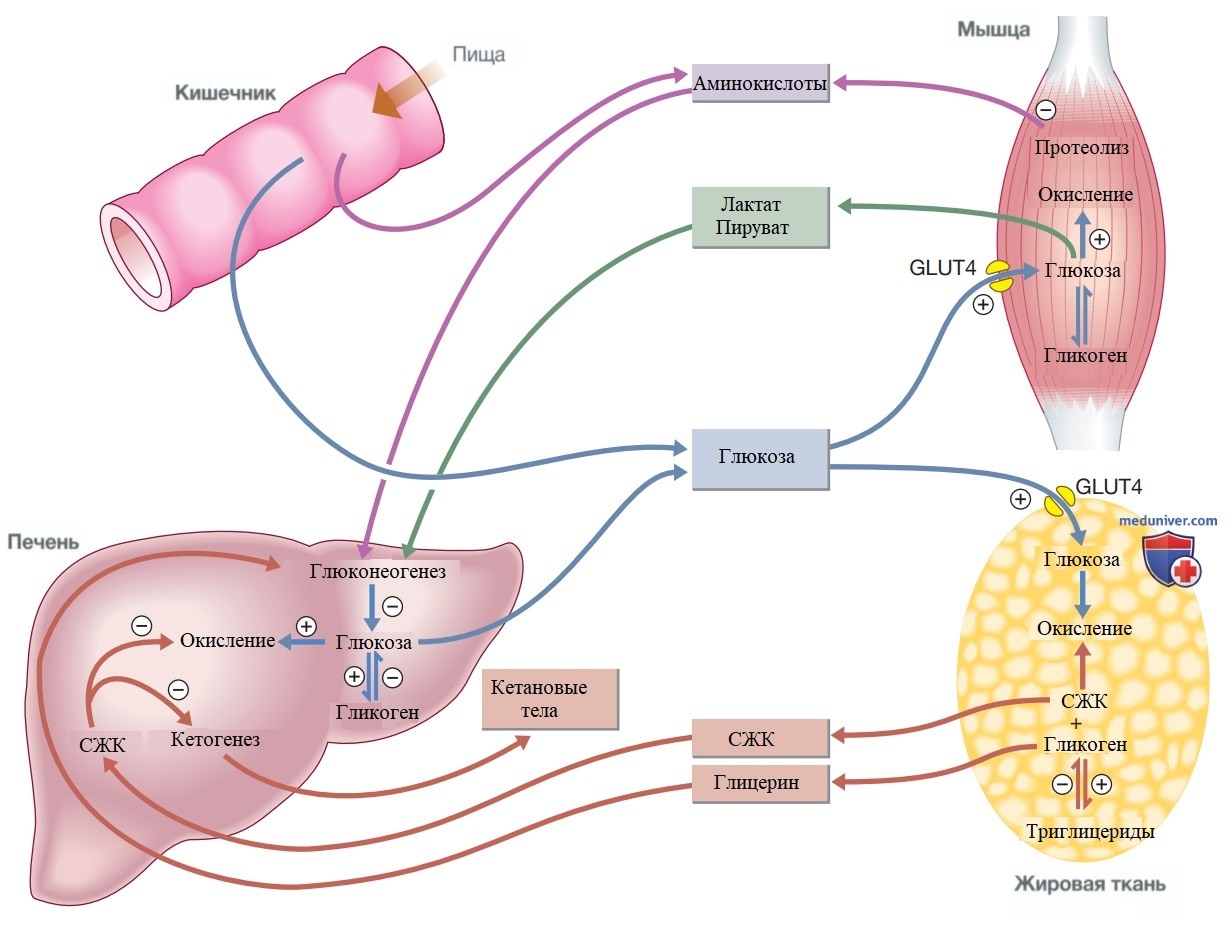
• стимуляции захвата глюкозы периферическими тканями (рис. 4).
Рисунок 4. Основные метаболические пути источников энергии и эффекты инсулина. Значок + обозначает стимуляцию, а - — супрессию инсулином. В ответ на повышение уровня глюкозы в крови (например, после приема пищи) высвобождается инсулин, подавляющий глюконеогенез и способствующий синтезу и накоплению гликогена. Инсулин усиливает захват глюкозы периферическими тканями, особенно в скелетных мышцах, и способствует ее накоплению (в форме мышечного гликогена). Он также стимулирует синтез белка и липогенез и подавляет липолиз. Инсулин управляет процессами высвобождения промежуточных метаболитов, включая аминокислоты (глутамин, аланин), трикарбоновые промежуточные продукты при окислении (лактат, пируват) и свободные жирные кислоты. При отсутствии инсулина, например, в период голодания происходят обратные процессы, которые способствуют глюконеогенезу в печени из гликогена, глицерина, аминокислот и других трикарбоновых предшественников
Постпрандиальное повышение уровня инсулина и глюкозы в системе воротной вены наряду со снижением концентрации глюкагона подавляет образование глюкозы в печени и приводит к преобладанию захвата глюкозы печенью. В зависимости от величины углеводной нагрузки в печени остается от 1/4 до 1/3 содержащейся в пище глюкозы. Кроме того, инсулин стимулирует захват глюкозы в скелетных мышцах и жировой ткани, опосредованный переносчиком глюкозы GLUT4.
Когда всасывание глюкозы из кишечника между приемами пищи снижается, концентрации инсулина и глюкозы в воротной вене падают, а уровень глюкагона повышается. Это приводит к увеличению образования глюкозы печенью за счет глюконеогенеза и расщепления гликогена. Теперь печень возобновляет чистое производство глюкозы, и гомеостаз глюкозы сохраняется. Основными субстратами для глюконеогенеза являются глицерин и аминокислоты, как показано на рис. 4.
г) Жировой обмен. Адипоциты (и печень) синтезируют триглицериды из неэтерифицированных СЖК и глицерина. Инсулин является основным регулятором не только метаболизма глюкозы, но и метаболизма жирных кислот. Высокий уровень инсулина после приема пищи способствует накоплению триглицеридов. Напротив, низкие уровни инсулина натощак позволяют осуществлять липолиз и высвобождение в кровоток СЖК (и глицерина), которые могут окисляться многими тканями. Их частичное окисление в печени обеспечивает энергию для управления глюконеогенезом, а также приводит к образованию кетоновых тел (ацетоацетат, который может восстанавливаться до β-гидроксибутирата или декарбоксилироваться до ацетона) в митохондриях гепатоцитов.
Кетоновые тела представляют собой органические кислоты, которые при образовании в небольших количествах окисляются и используются в качестве метаболического источника энергии.
Однако скорость утилизации кетоновых тел периферическими тканями ограниченна, и когда скорость образования в печени превышает скорость их удаления, возникает гиперкетонемия. Это физиологически происходит в период голодания, когда низкие уровни инсулина и высокие уровни катехоламинов увеличивают липолиз и доставку СЖК в печень.
Видео физиология гормонов поджелудочной железы и регуляции уровня глюкозы - профессор, д.м.н. П.Е. Умрюхин