Перирадикулярные ткани включают цемент, периодонтальную связку и костную ткань. Цемент — это минерализованная, аваскулярная соединительная ткань, состоящая из трех различных типов. Бесклеточный безволокнистый цемент покрывает зуб на протяжении всего ЦЭС. Расположение бесклеточного внешнего волокнистого цемента ограничено корональной половиной корня. Клеточный внутренний волокнистый цемент располагается на апикальной поверхности корня, где бесклеточный внешний волокнистый цемент не был отложен.
В матриксе цемента содержится множество факторов роста, таких как ИФР-1, факторы роста фибробластов (fibroblast growth factors, FGF), EGF, BMP, ТФР-β и фактор роста тромбоцитов (platelet-derived growth factor, PDGF).
Данные факторы роста в некоторых условиях могут высвобождаться; была показана их связь с пролиферацией, миграцией и дифференцировкой цементобластов в процессе восстановления цемента.
Периодонтальная связка представляет собой мягкую, специализированную соединительную ткань, которая связывает цемент с альвеолярной костью. Периодонтальная связка состоит из гетерогенных клеточных популяций и внеклеточного матрикса. Клетки периодонтальной связки включают остеобласты, остеокласты, фибробласты, эпителиальные клеточные островки Малассе, макрофаги, цементобласты и недифференцированные мезенхимальные клетки (стволовые клетки).
Фибробласты, остеобласты и эпителиальные клетки являются дифференцированными клетками, которые сохранили способность подвергаться ограниченному клеточному делению и пролиферации при их стимуляции соответствующими сигналами.
Мультипотентные MSC периодонтальной связки способны дифференцироваться в цементобластподобные клетки и клетки периодонтальной связки, а также в остеобласты. Внеклеточный матрикс периодонтальной связки состоит из коллагеновых волокон, фибронектина, эластина, других неколлагеновых белков и протеогликанов. Внеклеточный матрикс действует как слой для клеточной адгезии и способствует распространению клеток и цитоскелет-ной организации. Коллагеновые волокна (шарпеевское волокно) периодонтальной связки соединяют зуб с альвеолярной костью.
Периодонтальная связка сильно васкуляризирована и иннервирована. Ткань, расположенная апикальнее дентинно-цементного соединения, должна рассматриваться как часть периодонтальной связки, поскольку цемент не является нормальным компонентом ткани пульпы.
Эпителиальные клеточные островки Малассе (ЭКОМ) являются остатками гертвиговского эпителиального корневого влагалища, которое подвергается дезинтеграции после развития зуба, и присутствуют в периодонтальной связке рядом с поверхностью корня у всех зубов после формирования их корня. ЭКОМ в периодонтальной связке в норме находятся в состоянии покоя, но в случае апикального периодонтита они могут подвергаться стимуляции с дальнейшей пролиферацией. Считается, что они являются клеточным источником, который при правильной стимуляции может образовывать радикулярные кисты в некоторых случаях апикальных периодонтитов.
Альвеолярная кость, или альвеолярный отросток, — это часть кости челюстей, содержащая лунки для зубов. Альвеолярная кость состоит из внешней кортикальной пластинки, центральной пористой или губчатой кости и кости, выстилающей поверхность лунок. Костный матрикс содержит ИФР, ТФР-β, BMP, FGF и PDGF. Данные факторы роста необходимы для пролиферации, миграции и дифференцировки клеток-предшественников остеобластов в процессе заживления костной ткани.
Ответ перирадикулярных тканей на различные повреждения аналогичен таковому у других соединительных тканей в каких-либо участках организма. Ответ проявляется в виде воспалительной реакции, регулирующейся врожденным и адаптивным иммунными механизмами. Несмотря на то что микробная инфекция пульпы в корневых каналах является главной причиной апикального периодонтита, патологические изменения ПА-тканей при апикальном периодонтите обычно не вызываются собственно микробами, а в большей степени их токсинами, ядовитыми метаболическими побочными продуктами и дезинтегрированной тканью пульпы в системе корневых каналов.
Раздражители способны индуцировать как врожденный, так и адаптивный иммунный ответ; они могут либо активировать неантигенные пути, либо служить как антигены и активировать адаптивные ответы. Последующие воспалительные ответы различаются и могут включать изменения микроциркуляции, трансмиграции кроветворных клеток и плазматических белков из кровяного русла в тканевое пространство и активацию чувствительных нервов. Кроме того, эндотелиальные клетки, тучные клетки, тромбоциты, фибробласты, нейтрофилы, макрофаги, дендритные клетки, клетки врожденного и адаптивного иммунного ответа, иммуноглобулины, воспалительные медиаторы, провоспалительные цитокины, хемокины и нейропептиды также вовлекаются в воспалительный ответ.
Апикальный периодонтит может быть защитным или деструктивным в зависимости от динамического взаимодействия между микробной атакой и механизмами защиты организма-хозяина в ПА-тканях. К сожалению, бактериальная биопленка, образованная в системе корневых каналов с некротизированной пульпой, защищена от защитных механизмов организма-хозяина и антибиотикотерапии вследствие недостаточного количества сосудов микроциркуляции в системе корневых каналов. Следовательно, любые попытки пораженных ПА-тканей к репарации/регене-рации тщетны, поскольку бактериальные токсины и вредоносные побочные продукты метаболизма системы корневых каналов непрерывно выходят в ПА-область и раздражают ПА-ткани.
Появляющиеся доказательства говорят о том, что в большинстве случаев бактериальные биопленки в комплексе системы корневых каналов могут быть значительно уменьшены, но не уничтожены с помощью традиционных эндодонтических манипуляций, таких как механическая инструментальная обработка, антисептическая ирригация и интраканальная медикация. В том случае, если микробы системы корневых каналов эффективно уничтожаются или закрываются материалами для пломбировки корневых каналов и система корневых каналов надлежащим образом запечатана и защищена от коронального микроподтекания, тогда перирадикулярные ткани имеют возможность восстановления их истинного строения с помощью репаративных или регенеративных процессов.
Тем не менее наличие апикального периодонтита после лечения может объясняться стойкими микробными биопленками, и это знание стало поводом для начала крупных исследований по устранению биопленок.
а) Распространенность. Согласно эпидемиологическим исследованиям апикального периодонтита, его распространенность варьирует среди пациентов в возрасте от 20 до 30 лет (распространенность апикального периодонтита 33%), от 30 до 40 лет (40%), от 40 до 50 (48%), от 50 до 60 (57%) и старше 60 лет (62%). Большинство исследований распространенности апикального периодонтита проводится среди европейских и скандинавских стран. Согласно опросам ADA в 1990 г., оцениваемые 14 млн случаев лечения корневых каналов были выполнены только в США. Апикальный периодонтит является распространенной проблемой в сфере здоровья.
б) Этиология. Этиология, патогенез и гистопатология апикального периодонтита аналогичны таковым у маргинального пародонтита. Оба заболевания вызываются бактериальной инфекцией и включают патологические изменения альвеолярной кости, периодонтальной связки и цемента. Маргинальный пародонтит поражает коронально расположенные ткани пародонта, в то время как апикальный периодонтит действует на апикально расположенные ткани. Потеря кости является одной из характерных особенностей обоих заболеваний: при маргинальном пародонтите теряется кость гребня, а при апикальном периодонтите подвергается резорбции апикальная кость.
Апикальный периодонтит может быть вызван и экзогенными, и эндогенными факторами. Экзогенные факторы включают микробы и их токсины и вредные метаболические побочные продукты, химические агенты, механическое раздражение, чужеродные тела и травму. Эндогенные факторы включают метаболические продукты хозяина, такие как уратные и холестериновые кристаллы, а также цитокины или другие воспалительные медиаторы, которые активируют остеокласты. Эти раздражители могут активировать неантигенные или антигенные пути для запуска врожденных или адаптивных иммуновоспалительных ответов соответственно.
В системе корневых каналов первичной причиной апикальных периодонтитов является инфицирование пульпарной ткани, вызванное кариесом или другими способами. Классическое исследование Kakehashi и соавт. показало, что некроз пульпы и перирадикулярное воспаление у крыс в условиях опыта развились, когда пульпа зубов была подвержена действию микроорганизмов полости рта. Однако у стерильных лабораторных крыс некроз пульпы и перирадикулярное воспаление не возникли даже тогда, когда пульпа зубов была подвержена действию среды полости рта и закрыта стерильными остатками пищи. Аналогичный ответ наблюдался у людей.
С использованием бактериального культивирования было показано, что травмированные зубы у людей с интактными коронками и некротизированной пульпой без бактериальной контаминации не показали рентгенологического доказательства деструкции ПА-кости. Напротив, если бактерии были выделены из травмированных зубов с интактными коронками и некротизированной пульпой, то наблюдались рентгенологические свидетельства деструкции перирадикулярной кости. Эти важные открытия были повторены в нечеловеческих экспериментах над приматами. Когда пульпа интактных жизнеспособных зубов была намеренно девитализирована в асептических условиях и была оставлена в корневых каналах под непроницаемой для бактерий герметизирующей коронковой реставрацией на срок от 6 мес до 1 года, не наблюдалось никакой перирадикулярной воспалительной реакции.
Обобщая сказанное, существуют значительные доказательства того, что бактерии являются главным этиологическим фактором в развитии апикального периодонтита.
Бактериальные токсины (например, ЛПС, липотейхоевая кислота) и вредоносные метаболические побочные продукты, которые выходят из системы корневых каналов в ПА-ткани, способны индуцировать ПА-воспалительную реакцию. Эти вещества могут активировать врожденную иммунную систему посредством рецепторов, которые распознают стереотипные патоген-асссоциированные молекулярные паттерны (РАМР), обнаруженные в структуре этих токсинов. Различные классы микробов выделяют различные паттерны, которые распознаются различными PRR или TLR на клетках организма-хозяина, таких как фагоциты, дендритные клетки и В-лимфоциты. PRR или TLR закодированы в зародыше.
У млекопитающих существуют по крайней мере 10 видов TLR, каждый из которых, по-видимому, имеет отдельную функцию во врожденном иммунном распознавании. Например, ЛПС может стимулировать чувствительные нервные волокна для высвобождения CGRP и субстанции Р (SP) для развития вазодилатации и увеличения сосудистой проницаемости. ЛПС и липопротеины также могут активировать TLR на дендритных клетках для стимуляции дифференцировки Т-лимфоцитов. Некоторые подтипы TLR распознают общие структурные особенности различных токсинов (т.е. РАМР). Поскольку TLR синтезируются до инфицирования, они относятся к врожденной иммунной системе.
Апикальный периодонтит может вызывать как проникновение в ПА-ткани бактериальных токсинов, ферментов и вредоносных метаболических побочных продуктов, так и прямой инвазией в ПА-ткани микробов из системы корневого канала. Важно проводить различие между апикальным воспалением и апикальной инфекцией. Апикальное воспаление — это реакция ПА-ткани на раздражители, выходящие из системы корневого канала, которая проявляется вазодилатацией, повышенной сосудистой проницаемостью и экссудацией. Напротив, апикальная инфекция возникает из-за физического наличия патогенных микроорганизмов в ПА-тканях, которые постепенно вызывают разрушение ткани.
Возможно существование инфекции без воспаления, например, у пациента с ослабленным иммунитетом. Также может быть воспаление без инфекции, например, при инфаркте миокарда, инфаркте головного мозга и физическом или химическом повреждении. При вызванных инфекцией заболеваниях бактерии обычно присутствуют в вовлеченных в процесс тканях или органах, таких как острый некротический гингивит, маргинальный пародонтит, актиномикоз, туберкулез и бактериальный бронхит. Несмотря на то что апикальный периодонтит является прежде всего инфекционным заболеванием, бактерии обычно присутствуют не в ПА-тканях, а в системе корневых каналов, за исключением некоторых случаев апикального периодонтита, связанных с образованием абсцесса, или с создающим отток свищевым ходом , или экстрарадикулярной эндодонтической инфекцией.
Одна из основных текущих гипотез заключается в том, что апикальный периодонтит запускается путем проникновения в ПА-ткани бактериальных токсинов, ферментов и вредных метаболических побочных продуктов. Простое присутствие бактерий (колонизация) в некоторых апикальных периодонтитах не обязательно означает перирадикулярную инфекцию. ПА-инфекция связана как с вирулентностью, так и с числом и специфическими комбинациями микроорганизмов в ПА-тканях. Бактерии могут временно присутствовать в воспаленных перирадикулярных тканях только для того, чтобы быть уничтоженными защитными механизмами хозяина, тогда как очаг инфекции в системе корневых каналов эффективно устраняется механической обработкой, антисептической ирригацией и внутриканальными повязками.
Например, большинство апикальных периодонтитов с образованием абсцесса или свищевого хода хорошо заживают после нехирургического лечения корневого канала без необходимости системной антибиотикотерапии.
Первичная эндодонтическая инфекция в необработанных корневых каналах представляет собой полимикробную смесь с приблизительно равными пропорциями грамположительных и грамотрицательных видов, среди которых преобладают облигатные анаэробы. В зубах с запломбированными каналами с апикальным периодонтитом грамположительные микроорганизмы с относительно равным распределением факультативных и анаэробных видов, по-видимому, доминируют над другими микроорганизмами. Высокая распространенность Е. faecalis часто наблюдается в запломбированных корневых каналах, связанных с персистирующим апикальным периодонтитом. Эти вопросы более подробно описаны в отдельной статье на сайте - просим пользоваться формой поиска выше.
Физические (чрезмерная инструментальная обработка и избыточное пломбирование) и химические (ирриганты, внутриканальные повязки, материалы для пломбирования корневых каналов) поражения, а также травматическое повреждение ПА-тканей также могут вызывать апикальный периодонтит в зависимости от тяжести травмы и цитотоксичности химических веществ. Было показано, что такие инородные тела, как материалы для пломбирования корневых каналов, вызывают персистирующее ПА-воспаление. Однако возможность бактериальной контаминации в апикальных периодонтальных повреждениях, индуцированных чужеродными телами, не была тщательно исключена многими исследованиями, поэтому вероятно, что инородные тела служат переносчиками микроорганизмов.
Кроме того, инородные тела имеют необычное свойство поддерживать инфекцию, поскольку они могут снизить инфекционную составляющую бактерий (LL) и вызывать развитие фагоцитарного дефекта или потерю защищенности посредством действия на гранулоциты (LL, ОК).
И хотя большинство материалов для пломбирования корневых каналов не являются инертными и в определенной степени могут вызывать воспаление, в целом они биосовместимы и хорошо переносятся ПА-тканями.
Гистологически было показано, что заболевания пародонта может вызывать воспалительные заболевания пульпы и ПА-тканей.
б) Инфекция как конфликт между хозяином и паразитами. Каждый инфекционный процесс является состязанием между способностью микроорганизма размножаться, распространяться и вызывать заболевание и возможностью организма-хозяина контролировать и в конечном счете ликвидировать микроорганизм. У хозяина есть физические барьеры — поверхностный эпителий, эмаль и дентин, а также врожденная и адаптивная иммунная защита для предотвращения пульпарной и ПА-инфекции. Тем не менее паразиты тоже обладают оружием, что приводит к ингибированию фагоцитоза, ингибированной лизосомальной функции, снижению эффективности уничтожения фагоцитами, инактивации системы комплемента и иммуноглобулинов, а также специфическими механизмами, которые позволяют проникать через физические барьеры хозяина.
Инфицирование ткани может проявляться различными гистопатологическими особенностями в результате специфических взаимодействий хозяин-паразит. Многие инфекции бессимптомны у более чем 90% людей. Например, некроз пульпы и хронический апикальный периодонтит, вызванный инфекцией корневого канала, обычно бессимптомны, и пациенты часто удивляются, обнаружив, что эта инфекция присутствовала в течение времени, достаточного для возникновения деструкции ПА-кости. Следовательно, нет простой взаимосвязи между инфекцией и клиническими симптомами апикального периодонтита, за исключением случаев симптоматического апикального периодонтита и острого апикального абсцесса.
в) Патогенез. Когда пульпа инфицирована/воспалена, многие врожденные и адаптивные иммунные клетки высвобождают повышенное число различных воспалительных медиаторов, включая цитокины, хемокины и нейропептиды. По мере распространения пульпарного воспаления воспалительные медиаторы начинают изменять физиологию ПА-тканей. Клинически наблюдаемые изменения при рентгенографическом обследовании — это расширение пространства периодонтальной связки или развитие апикальных остеолитических поражений вследствие резорбции кости. Потеря кости в основном вызвана активированными остеокластами. Обнаружено, что многие цитокины, такие как IL-1, IL-11, IL-17 и ФНО-α, способны индуцировать дифференцировку и активацию клеток-предшественников остеокластов. Резорбция кости в ПА-тканях, вызванная воспалением, сопровождается захватом иммунных клеток, которые в сущности строят оборонительную линию против распространения микробной инвазии из корневого канала.
Патогенез апикального пародонтита включает врожденные и адаптивные иммунные реакции, а также ответную реакцию чувствительного нерва в ПА-тканях. Иммунные клетки, присутствующие в перирадикулярных поражениях человека, состоят из лимфоцитов, макрофагов, плазматических клеток, нейтрофилов, дендритных клеток и натуральных киллеров (NK), в основном с первыми двумя типами. Характерные особенности врожденного и адаптивного иммунитета приведены в табл. 1.
1. Врожденный иммунный ответ:
1.1 Специфичность врожденного иммунного ответа. В последние годы понятие неспецифической природы врожденного иммунитета изменилось после обнаружения сети закодированных в зародыше рецепторов, PRR, упомянутых ранее, которые распознают специфические молекулярные паттерны микроорганизмов. PRR могут экспрессироваться на поверхности клетки (макрофаги, дендритные клетки, нейтрофилы, NK-клетки, В-клетки), во внутриклеточных компартментах или секретироваться в крови и тканевых жидкостях. Существуют многочисленные микробные конститутивные и скрытые продукты, такие как РАМР, что также было отмечено ранее. Важно отметить, что PRR врожденной иммунной системы распознают РАМР.
Специфичность врожденного иммунитета связана с распознаванием РАМР микроорганизмов с помощью PRR, таких как TLR, большинства клеток организма-хозяина. Активация PRR вызывает многочисленные ответы хозяина, включая опсонизацию, активацию комплемента и коагуляционных каскадов, фагоцитоз, активацию провоспалительных сигнальных путей и индукцию апоптоза. Например, TLR4/CD14 представляет собой рецептор для грамотрицательных бактериальных ЛПС. Мыши с TLR4-мутированным СЗНО/ HeJ (сниженный ответ на ЛПС) имели пониженный ответ на грамотрицательные бактерии и были сильно восприимчивы к инфекциям, вызванным Salmonella Typhimurium или Neisseria meningitidis.
Важно отметить, что существует пониженная экспрессия IL-1 и IL-12 и уменьшение перирадикулярной деструкции кости у TLR-4-дефицитных мышей, когда зубы подвергаются воздействию на пульпу и инфицированию смесью 4 анаэробных патогенов: Prevotella intermedia, Fusobacterium nucleatum, Streptococcus intermedius (G+) и Peptostreptococcus micros (G+). Кроме того, показано, что ЛПС способен вызывать боль с помощью прямой активации TLR4/CD14, которые экспрессируются на ноцицептивных чувствительных нейронах. Таким образом, рецептор TLR4 PRR является важным звеном в развитии одонтогенных инфекций.
Липотейхоевая кислота клеточной стенки грамположительных бактерий также может стимулировать врожденный иммунитет по аналогичному ЛПС механизму. TLR2 играет важную роль в обнаружении грамположительных бактерий и участвует в распознавании различных микробных компонентов, в том числе липотейхоевой кислоты, липопротеинов и пептидогликана. Важность TLR2 в защите организма-хозяина против грамположительных бактерий была продемонстрирована с использованием TLR2-дефицитных (TLR2-/-) мышей, которые, как было установлено, в высшей степени восприимчивы к внедрению Staphylococcus aureus или Streptococcus pneumoniae. Липотейхоевая кислота также стимулирует выделение лейкоцитами медиаторов воспаления, включая ФНО-α, IL-1β, IL-6, IL-8 и простагландин (ПГ) Е2, которые, как известно, играют важную роль в различных фазах воспалительного ответа.
Все эти медиаторы воспаления были обнаружены в образцах ПА-ткани, и каждый из них имеет хорошо известный эффект разрушения ткани в результате активации различных ответов хозяина.
Врожденный иммунный ответ на бактериальную инфекцию индуцирует выделение провоспалительных цитокинов, хемокинов и костимуляторов, которые необходимы для активации и проявления действия адаптивного иммунного ответа. Врожденная иммунная система способна распознавать чужеродные и аутоантигены, в то время как адаптивная иммунная система этой способностью не обладает. Таким образом, многие аутоиммунные заболевания являются нарушением адаптивного иммунитета.
1.2 Неспецифический врожденный иммунный ответ. Механизм первичной неспецифической врожденной иммунной защиты при апикальном периодонтите состоит в фагоцитозе микробов специализированными фагоцитами, такими как полиморфно-ядерные лейкоциты (ПМЯЛ) и макрофаги. Тканевое воспаление приводит к захвату ПМЯЛ из кровотока в перирадикулярную ткань. Активированные ПМЯЛ вызывают резкое увеличение потребления кислорода, хорошо известное как респираторный взрыв, приводя к высвобождению кислородных радикалов, семейству крайне разрушительных короткоживущих веществ, разрушающих расположенные поблизости микроорганизмы и клетки хозяина. Фагоцитированные микробы или инородные частицы подвергаются действию токсичной среды, содержащей специфические и азурофильные гранулы и свободные кислородные радикалы, и в конечном счете распадаются. ПМЯЛ также обладают внеклеточным механизмом уничтожения посредством нейтрофильных внеклеточных ловушек, которые являются внеклеточными структурами, состоящими из хроматина со специфическими белками из нейтрофильных гранул.
После активации (например, IL-8, ЛПС, бактериями, грибами, активированными тромбоцитами) нейтрофилы запускают клеточную программу, называемую апоптозом, что приводит к их гибели и формированию нейтрофильных внеклеточных ловушек, обладающих антимикробной активностью. Кроме того, их роль во врожденном иммунитете как профессиональных фагоцитов, макрофагов также заключается в работе в качестве АПК, экспрессирующих МНС (главный комплекс гистосовместимости) молекул класса II, которые взаимодействуют с антиген-специфичными клонами Т-хелперов. Циркулирующие моноциты являются предшественниками как тканевых макрофагов, так и множества подтипов дендритных клеток. Подробности иммунологической активности нейтрофилов и макрофагов в патогенезе ПА-поражения описаны в отдельных статьях на сайте - просим пользоваться формой поиска выше.
2. Адаптивный/специфический иммунный ответ. Специфичность адаптивного иммунитета регулируется на генетическом уровне в В- и Т-лимфоцитах сложным процессом, ведущим к образованию молекул, которые распознают и связываются с чужеродными или аутоантигенами. Эти молекулы являются специфическими рецепторами Т-клеток (Т-клеточные рецепторы к антигену, TCR), а также В-клеток (В-клеточные рецепторы к антигену, BCR, также называемые иммуноглобулинами). TCR на Т-клетках взаимодействуют с антигенами, которые представлены с помощью АПК, экспрессирующих молекулы МНС вместе с другими вспомогательными молекулами, тогда как BCR на В-клетках взаимодействуют с антигенами напрямую. BCR может секретироваться в кровоток или в тканях в виде антител. Вариабельный фрагмент обоих TCR и BCR белков перестраивается на геномном уровне с помощью генетической рекомбинации V(D)J сегментов. Согласно оценкам, общее многообразие после этой рекомбинации для TCR составляет приблизительно 1021 и для BCR составляет приблизительно 1016, при этом генерируется набор различных индивидуальных Т- и В-клеточных клонов.
Каждый клон Т- или В-клеток, образованный в костном мозге, несет определенные TCR и BCR. Они подвергаются процессу положительного и отрицательного отбора, через который большинство клонов удаляется с помощью апоптоза, поскольку они связываются с аутоантигенами. Этот начальный процесс «отрицательного скрининга» снижает вероятность аутоиммунных расстройств. Только те клоны, которые не взаимодействуют с аутоантигенами, высвобождаются в лимфатическую систему и систему кровообращения. Наивные Т-клетки циркулируют туда и обратно между лимфатической системой и кровяным руслом, пока они не сталкиваются с чужеродными антигенами на АПК. Около 97% Т-клеток подвергается апоптозу, и лишь небольшой их процент транспортируется на периферию как зрелые Т-клетки.
Взаимодействие между TCR и комплексом антигенный пептид/МНС и ко-стимуляторами активирует Т-клетки, что приводит к синтезу фактора роста Т-клеток, IL-2 и его рецептора, который вызывает Т-клеточную клональную экспансию/пролиферацию. Некоторые из этих Т-клеток дифференцируются в усиленные, эффекторные Т-клетки, а другие становятся клетками памяти. Существует несколько Т-клеточных субпопуляций, классифицированных по выполняемой ими функции: 1) Т-клетки-хелперы (TH), 2) Т-регуляторные клетки (Treg), 3) клетки Т-супрессоры (TS) и 4) цитотоксическая (цитолитическая) (ТС) Т-клетка. Некоторые из них можно отличить по маркерам на их клеточной поверхности, цитокиновому профилю или факторам транскрипции. Для дополнительных подробностей см. также главу 13.
Под антигенной стимуляцией наивные CD4 Т-клетки пролиферируют и дифференцируются в Th0, которые постепенно претерпевают развитие в Th1- или Th2-клетки. Моноцитоидная дендритная клетка (DC1) индуцирует ответы Th1-типа. Плазмацитоидная дендритная клетка (DC2) избирательно индуцирует Th2-ответы. Каждый подтип Th-клеток имеет определенные функции и набор цитокинов. Th1-клетки главным образом выделяют IL-2 и ИФН-γ, которые активируют макрофаги и индуцируют выработку опсонизирующих антител В-клетками. Th2-клетки вырабатывают IL-4, -5, -10 и -13, которые активируют В-клетки с целью выработки нейтрализующих антител. Th0 также может перейти в Th17 в результате активации IL-6 и ТФР-β и вырабатывать IL-17, мощный провоспалительный цитокин. В целом, ТЫ и Th2 имеют взаимно ингибирующий эффект. Развитие CD4 Th-клеток включает столкновение антигена на АПК в комплексе с МНС класса II.
Все клетки выделяют МНС класса I, но только некоторые клетки экспрессируют МНС класса II. Эти клетки, вырабатывающие МНС класса II, образуют популяцию АПК в организме и включают: 1) дендритные клетки, 2) макрофаги, 3) В-клетки, 4) эндотелиальные клетки сосудов, и 5) эпителиальные клетки. Первые 3 называют профессиональными АПК, поскольку они предназначены для этой функции. Две последние АПК в нормальных условиях находятся в состоянии покоя, но возможна их активация с выделением МНС класса II при воздействии повышенной концентрации ИФН-γ.
Дендритные клетки и макрофаги фагоцитируют чужеродные антигены, тогда как В-клетки используют связанный с мембраной иммуноглобулин для присоединения и поглощения антигенов. Другие АПК подвергают эндоцитозу в цитоплазму чужеродные антигены для их передачи. Переданные антигены частично распадаются на маленькие пептиды. Многие из них имеют длину от 10 до 30 аминокислот и способны к присоединению к новообразованным молекулам МНС класса II до того, как комплекс антиген/МНС класса II транспортируется на поверхность клеток и будет представлен TCR CD4+ Т-клеток.
Противоречивые свидетельства говорят о том, что CD8+Ts и CD8+CTL представляют собой различные субпопуляции CD8+ Т-клеток. Ts являются ограниченными МНС класса I CD8+/CD28-Ts-клетками, которые действуют на АПК контакт-зависимым образом, делая их толерогенными к Th-клеткам. Они тормозят пролиферацию Th-клеток. Цитотоксические Т-клетки (CD8+Tc), также известные как цитолитические Т-лимфоциты (CTL), являются подтипом Т-клеток, который убивает клетки мишени, экспрессирующие МНС-ассоциированные пептидные антигены. Большинство Т-клеток выделяют CD8 и распознают антигены, распавшиеся в цитозоли и выведенные на поверхность клетки в комплексе с молекулами МНС класса I клеток-мишеней. Функционирующие Т-клетки требуют специфических мембраносвязанных цитоплазматических гранул, включающих мембранные порообразующие белки, называемые перфорином или цитолизином, и ферменты, называемые гранзимами.
Роль В-клеток в адаптивном иммунитете главным образом заключается в выработке антител, которые составляют гуморальный ответ организма. Рекомбинация гена V(D)J возникает и в тяжелой, и в легкой (κ и λ) цепях. Рекомбиназная система, которая является ферментативным комплексом, состоящим из нескольких ферментов — включая гены 1 и 2, активирующие рекомбинацию (RAG-1, RAG-2), терминальную дезоксинуклеотидилтрансферазу (англ, terminal deoxynucleotidyl transferase, TdT), ДНК-лигазу IV, Ku-белки и XRCC4, — необходима для успешной рекомбинации. Эта рекомбинантная система также используется для рекомбинации TCR. Зрелые В-клетки, коэкспрессирующие IgM/IgD, подвергаются переключению изотипа посредством процесса, называемого свич-рекомбинацией, после встречи с антигеном. Реорганизованный сегмент гена V(D)J рекомбинируется с С-регионом гена (γ, ε или α), регулирующим последующие звенья сигнальных каскадов, и удаляется промежуточная последовательность ДНК. Это дает начало другим классам иммуноглобулинов (IgG, IgE, IgA), помимо IgM.
Кроме того, для переключения изотипа активированные Т-клетки также подвергаются соматической мутации в V регионе гена, приводя к аффинному созреванию антител или альтернативному сплайсингу VDJ РНК к мембранному или секретированному иммуноглобулину мРНК. Секретируется большое количество антител, когда В-клетки терминально дифференцируются в плазматические клетки. Способность антигенов избирательно стимулировать дифференцировку плазматических клеток говорит в пользу клинических данных о том, что плазматические клетки, выделенные из ПА-очагов поражений, секретируют специфические антитела против конкретных бактерий, обнаруженных в прилежащей системе корневых каналов.
Иммунный ответ и роль субпопуляций лимфоцитов в поражениях апикального периодонтита были исследованы с использованием лимфоцит-дефицитных грызунов в качестве модели для изучения. Т-клеточный дефицит, по-видимому, ускорял потерю кости в ранней фазе апикальных периодонтальных поражений (2 нед), но не влиял на общий курс развития поражения. Используя RAG-2 (активирующие рекомбинацию) SCID-мышей (Т- и В-клеточно-дефицитные), было установлено, что приблизительно у трети RAG-2-мышей развились эндодонтические абсцессы, тогда как ни у кого в иммунокомпетентной контрольной группе абсцессов не было. В другом исследовании специфичные мыши, лишенные RAG-2, были использованы для определения того, какие иммунные элементы были важны для защитного механизма при эндодонтической инфекции. Результаты продемонстрировали, что В-клетки, а не Т-клетки играли основную роль в предупреждении диссеминации эндодонтической инфекции. Следовательно, и Т-, и В-лимфоциты опосредуют наблюдаемые иммунные ответы в поражениях при апикальном периодонтите.
3. Нейрогенное воспаление. Некоторые первичные афферентные волокна в результате стимуляции разнообразными раздражителями высвобождают нейропептиды, которые вызывают вазодилатацию, экстравазацию белков и захват/регуляцию иммунных клеток, таких как макрофаги, нейтрофилы, тучные клетки и лимфоциты. Это явление называется нейрогенным воспалением. Основными нейропептидами в индукции нейрогенного воспаления является CGRP для вазодилатации и SP для индукции экстравазации белков. Нейропептиды и их рецепторы широко распространены по всему телу. Во время воспаления присутствует разрастание афферентных волокон и местные подъемы воспалительных медиаторов, которые запускают выделение нейропептида, ведущее к нейрогенному воспалению. Помимо главнейших функций ключевых нейропептидов, которые вызывают первый признак воспаления — вазодилатацию и повышение сосудистой проницаемости, роль этих нейропептидов в процессе воспаления является, как известно к настоящему времени, намного более сложной.
При развитии очагов хронического апикального периодонтита нейропептиды также вовлекаются в иммунную регуляцию, костную резорбцию и процесс заживления. В достаточных концентрациях SP повышает секрецию IL-1, ФНО-а и IL-6 макрофагами и стимулирует пролиферацию Т-лимфоцитов и усиливает антиген-индуцированную выработку ИФН-γ Т-клетками. Некоторые нейропептиды, такие как SP, вызывают повышающую регуляцию иммунных и воспалительных реакций, тогда как другие нейропептиды, такие как ВИП и нейропептид Y, ингибируют воспалительные ответы. Синергетические взаимодействия между нейропептидами, такими как CGRP и другие воспалительные медиаторы, эйкозаноиды и брадикинин, говорят о сложном взаимодействии этих молекул при иммунном ответе.
В хронических апикальных периодонтальных очагах поражения на некоторых иммунных клетках, включая макрофаги и лимфоциты, выделяются специфические рецепторы для SP и CGRP. И CGRP, и ВИП могут играть роль в торможении костной резорбции посредством супрессии функционирования остеокластов. Уровень ВИП в очагах апикального периодонтита обратно пропорционален размеру поражения. Изучения клеточной культуры остеокластов продемонстрировали, что наличие больших концентраций ВИП приводит к понижению их способности образовывать лакуны костной резорбции, вызывая быстрое сжатие цитоплазмы и сокращение клеточной подвижности. ВИП действует на макрофаги, вызывая блокирование образования ФНО-α, IL-6 и IL-12, что говорит о том, что ВИП может играть роль в контролировании роста апикальных периодонтальных поражений.
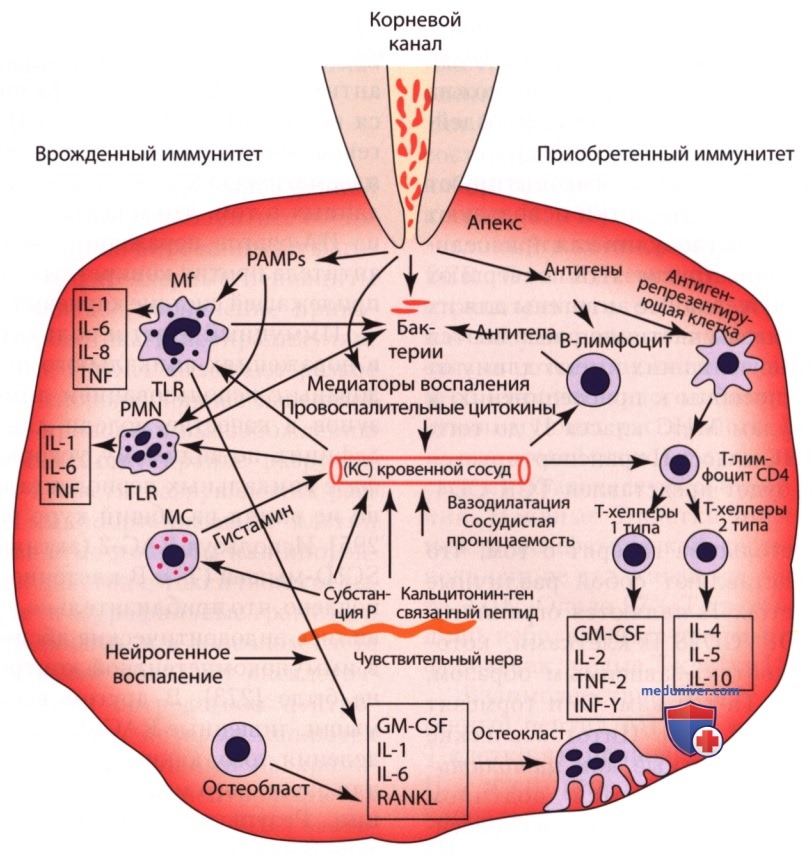
Основные врожденные и адаптивные иммунные реакции и нейрогенное воспаление в патогенезе апикального периодонтита вызваны инфицированием корневого канала и изображены на рис. выше.