а) Воспалительный ответ фагоцитов. Система фагоцитов состоит из гранулоцитов (нейтрофилы, эозинофилы и базофилы) и мононуклеарных фагоцитов (моноциты и тканевые макрофаги). Нейтрофилы и мононуклеарные фагоциты разделяют основные функции, в т.ч. характерные свойства крупных частиц — поглощение и уничтожение микробов. Фагоциты участвуют во врожденном иммунном ответе, но также помогают инициировать приобретенный иммунитет. Мононуклеарные фагоциты, включая тканевые макрофаги и циркулирующие моноциты, рассмотрены в отдельной статье на сайте - просим Вас пользоваться формой поиска по сайту выше.
Нейтрофилы — быстрое эффекторное звено врожденной иммунной системы. Они циркулируют в кровотоке 6 ч (табл. 1), но в ответ на специфические хемотаксические стимулы прикрепляются к эндотелию сосудов и трансмигрируют в ткани. Там они поглощают и уничтожают микробы, вырабатывают хемотаксические сигналы для привлечения большего количества нейтрофилов, дендритных клеток и др. инициаторов приобретенного иммунного ответа.
б) Гемопоэз. Систему гемопоэтических клеток-предшественников можно рассматривать как совокупность функциональных отделов. Элементарный отдел состоит из редких плюрипотентных стволовых клеток, обладающих высокой способностью к самообновлению и дающих начало более зрелым стволовым клеткам, в т.ч. клеткам лимфоидного и миелоидного ряда (рис. 1). Обычные лимфоидные клетки-предшественники дают начало предшественникам Т- и В-клеток и их зрелому потомству. Обычные миелоидные клетки-предшественники дают начало коммитированным однолинейным предшественникам дочерних клеток через случайный процесс поэтапного ограничения возможности развития клеточных линий.
Рисунок 1. Основные источники и области действия цитокинов, а также потребности в факторах транскрипции для гемопоэтических клеток. Клетки микросреды костного мозга, такие как макрофаги, эндотелиальные клетки и ретикулярные фибробластоидные клетки, продуцируют макрофагальные, гранулоцитарно-макрофагальные и гранулоцитарные колониестимулирующие факторы (М-КСФ, ГМ-КСФ, Г-КСФ), IL-6 и фактор стволовых клеток (ФСК, клеточные источники точно не определены) после индукции эндотоксином (макрофаги) или IL-1/ФНО (эндотелиальные клетки и фибробласты). Т-клетки продуцируют IL-3, ГМ-КСФ и IL-5 в ответ на АГн-стимуляцию и стимуляцию IL-1. Во время гемопоэтической дифференцировки эти цитокины обладают аналогичными функциями, и оптимальное развитие требует комбинации факторов раннего и позднего действия для всех линий. Факторы транскрипции, важные для выживания или самообновления стволовых клеток, отмечены красным вверху, а стадии гемопоэза, заблокированные после истощения указанных факторов транскрипции, отмечены красным для мультипотентных и коммитированных предшественников. BMP — костный морфогенетический протеин; Wnt — суперсемейство секретируемых гликопротеинов, участвующих в передаче сигналов между клетками; Jagged (JAG1) — мембранный белок, лиганд для рецептора Notch1; LTR-HSC — долгосрочные гемопоэтические стволовые клетки.
Способность линейно-специфичных коммитированных предшественников пролиферировать и дифференцироваться в соответствии с потребностью организма обеспечивает гематопоэтической системе известный диапазон ответа на меняющиеся требования к продукции зрелых клеток крови.
Пролиферация, дифференцировка и выживаемость незрелых гемопоэтических клеток-предшественников регулируются гемопоэтическими факторами роста, семейства гликопротеинов. Кроме регуляции пролиферации и дифференцировки предшественников, эти факторы влияют на выживание и функцию зрелых клеток крови. Во время гранулопоэза и монопоэза множественные цитокины регулируют клетки на каждой стадии дифференцировки от плюрипотентных стволовых клеток до неделящихся, зрелых дифференцированных клеток (моноцитов, нейтрофилов, эозинофилов и базофилов). По мере созревания клетки теряют рецепторы большинства цитокинов, особенно тех, которые влияют на раннее развитие клеток. При этом сохраняются рецепторы цитокинов, влияющие на их мобилизацию и функцию (напр., факторы, стимулирующие колонии гранулоцитов и макрофагов).
Зрелые фагоциты также экспрессируют рецепторы хемокинов, которые помогают направлять клетки к участкам воспаления. Хемокиновые рецепторы CXCR4 и его лиганд SDF-1 играют ключевую роль в удержании развивающихся миелоидных клеток в костном мозге.
в) Созревание и кинетика нейтрофилов. Процесс созревания интрамедуллярных гранулоцитов включает стадию изменения ядерной конфигурации и накопление специфических в/цитоплазматических гранул. Микроокружение костного мозга поддерживает постоянное обновление нейтрофилов периферической крови за счет экспрессии стромальными клетками факторов роста и дифференцировки. Факторы роста, такие как гранулоцитарный колониестимулирующий фактор и гранулоцитарно-макрофагальный колониестимулирующий фактор, не только стимулируют деление клеток, но и индуцируют экспрессию факторов транскрипции, которые регулируют биосинтез функциональных компонентов нейтрофилов (гранулированных белков). Фактор транскрипции PU.1 играет важную роль в процессе миелопоэза (как «+» регуляторный элемент и как супрессор GATA1 — фактора транскрипции, который управляет немиелоидной дифференцировкой).
Другие факторы транскрипции, такие как Runxl (AML1), c-myb, CDP, С/ЕВР α, С/ЕВРγ и MEF, экспрессируются в миелобластах и промиелоцитах, некоторые из них необходимы для экспрессии белка азурофильных гранул. Когда клетки переходят в стадию миелоцитов, Runxl и myb подавляются, а экспрессия PU.1 и С/ЕВРе повышается, чтобы инициировать терминальную дифференцировку.
Гранулоциты живут в кровотоке 6-12 ч, поэтому для поддержания уровня циркулирующих гранулоцитов 5х103 кл./мкл требуется ежедневная продукция 2х104 кл./мкл крови (табл. 1). Относительно небольшой пул периферической крови включает быстро меняющиеся циркулирующие и маргинальные пулы. Маргинальный пул обеспечивает вход в тканевую фазу, где нейтрофилы могут жить в течение нескольких часов или дней.
Циркулирующий пул насыщается и восполняется гораздо большей популяцией зрелых нейтрофилов и миелоидных предшественников, составляющих соответственно резерв костного мозга и пролиферирующих пулов. Процесс пролиферации миелоидных клеток, протекающий на протяжении 5-ти митотических делений, в миелобластах, промиелоцитах и миелоцитах происходит только на первых 3 стадиях развития нейтрофилов. После стадии миелоцитов клетки окончательно дифференцируются на неделящиеся, созревающие метамиелоциты, фиброзные волокна и нейтрофилы.
Созревание нейтрофилов приводит к ядерной конденсации и лобуляции, а также к последовательному образованию специфических популяций гранул. Миелобласт — в некоторой степени недифференцированная клетка с крупным овальным ядром, большим ядрышком и недостаточным количеством гранул. Промиелоциты получают пероксидаза-«+» азурофильные (первичные) гранулы, затем миелоциты и метамиелоциты — специфические (вторичные) гранулы, а третичные гранулы и секреторные везикулы развиваются на заключительной стадии созревания нейтрофилов.
г) Нейтрофильная функция. Реакция нейтрофилов инициируется, когда циркулирующие нейтрофилы, проходя через посткапиллярные венулы, обнаруживают низкие уровни хемокинов и др. хемотаксических в-в, образующихся на месте инфекции. Последовательность событий по мере того, как нейтрофилы циркулируют с кровотоком, встречают и уничтожают бактерии, контролируется серией биохимических процессов, сбои в которых вызваны генетическими нарушениями функций нейтрофилов (рис. 2). Понимание клеточной биологии фагоцитарной функции базируется на упомянутых нарушениях функций нейтрофилов.
Рисунок 2. Опосредованная нейтрофилами воспалительная реакция и связанные с ней синдромы дисфункции нейтрофилов. Циркулирующие нейтрофилы свободно прикрепляются к эндотелию через селектины и катятся по стенке сосуда, пока не достигнут места инфекции. Воспалительные монокины, IL-1 и ФНО активируют эндотелиальные клетки для экспрессии Е- и Р-селектинов. Е- и Р-селектины служат контррецепторами для сиалильных нейтрофилов Lewis X. Lewis X вызывают скатывание нейтрофилов с низкой авидностью. Активированные эндотелиальные клетки экспрессируют ICAM-1, который служит контррецептором для молекул нейтрофильного β-интегрина. Это приводит к распространению лейкоцитов с высокой авидностью и началу трансэндотелиальной миграции в месте инфекции. Нейтрофилы проникают через базальную мембрану сосудов с высвобождением протеаз и реактивных окислительных промежуточных продуктов, вызывая локальное разрушение окружающей ткани в местах высоких концентраций хемотаксических факторов, и мигрируют к месту инфекции, где они поглощают и убивают бактерии.
Подгруппы циркулирующих нейтрофилов слабо прикрепляются к сосудистому эндотелию, используя низкоаффинные рецепторы — селектины, они передвигаются по эндотелию (перекатываются) и образуют маргинальный пул. Растворимые эффекторы воспаления вызывают незначительные изменения в молекулах поверхностной адгезии на эндотелиальных клетках в месте инфицирования. Движение «катящихся» нейтрофилов обеспечивает более интенсивное воздействие на них активирующих факторов — ФНО или IL-1 (рис. 2). Воздействие на нейтрофилы тех же самых активирующих факторов вызывает качественные и количественные изменения в семействе рецепторов адгезии β2-интегрина (группа поверхностных молекул CD11/CD18), что приводит к плотной адгезии между нейтрофилами и эндотелиальными клетками в месте воспаления и к трансмиграции нейтрофилов в ткань.
Пройдя через эндотелий, нейтрофил под воздействием градиента хемокинов или др. хемоаттрактантов мигрирует к участкам инфекции. Миграция нейтрофилов — сложный процесс, включающий этапы взаимодействия с рецептором, передачу сигнала и ремоделирование актиновых микрофиламентов, частично составляющих цитоскелет. Полимеризация и деполимеризация актина происходит за 8-секундные циклы, запускает циклическое растяжение и сжатие богатой актином ламеллы передней части нейтрофила. Рецепторы на переднем крае ламеллы определяют градиент аттрактанта, следуют за микроорганизмами, затем поглощают и уничтожают их. Когда нейтрофил достигает места инфекции, он распознает патогены с помощью Fc-Ig и рецепторов комплемента, толл-подобных рецепторов, рецепторов фибронектина и других молекул адгезии.
Нейтрофилы поглощают микробы, покрытые опсонинами, белками сыворотки, такими как Ig, и компонентом комплемента C3. Патогены попадают в закрытую вакуоль, фагосому (рис. 3), в которой одновременно происходят две клеточные ответные реакции, необходимые для оптимальной микробицидной активности: дегрануляция и активация НАДФН-зависимой оксидазы. Слияние мембран нейтрофильных гранул с мембраной фагосомы доставляет в фагосому мощные противомикробные белки и небольшие пептиды.
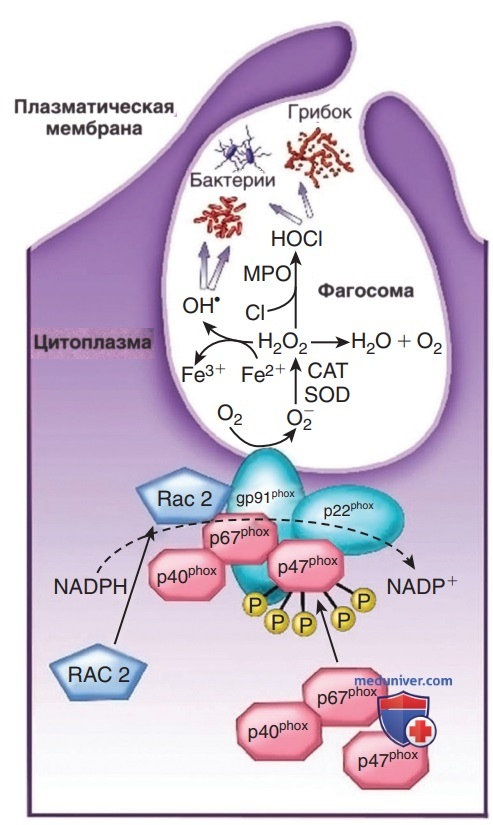
Рисунок 3. Компоненты и активация НАДФН-оксидазы. При активации фагоцитарных клеток 3 цитозольных компонента НАДФН-оксидазы (красный цвет) (р67 phox, р47 phox и р40 phox) + малый белок ГТФ (GTPase) Rac2 перемещаются на мембрану фагоцитарной вакуоли. Субъединица р47 phox связывается с мембранным компонентом флавоцитохрома b558 (сине-зеленый) НАДФН-оксидазы (gp91 phox + р22 phox). НАДФН-оксидаза катализирует образование супероксида, передавая электрон от НАДФН к молекулярному кислороду (О2), тем самым образуя супероксидный свободный радикал. Нестабильный супероксид-анион превращается в перекись водорода спонтанно или под действием супероксиддисмутазы (SOD). Н2О2 может следовать различным метаболическим путям с образованием более мощных реактивных окислителей, таких как ОН* или HOCl), или разлагаться до Н2О+О2.
Сборка и активация НАДФН-оксидазы также происходят на мембране фагосомы (рис. 3), генерируя большие количества супероксида (O2) из молекулярного кислорода, который разлагается с образованием пероксида водорода (Н2О2) и синглетов кислорода. Миелопероксидаза, главный компонент гранул азурофилов, катализирует реакцию Н2О2 с повсеместно присутствующими ионами хлорида для создания хлорноватистой кислоты (HOCl) в фагосоме. Н2О2 и HOCl — сильно действующие микробицидные агенты, которые разрушают и удаляют патогены из участков инфекции.
Кроме того, нейтрофилы секретируют ряд цитокинов и хемокинов, которые привлекают больше нейтрофилов для борьбы с инфекцией, также моноциты и макрофаги, которые обладают микробицидной функцией и функцией поглощения, способствуют представлению АГн, чтобы помочь инициировать адаптивный иммунный ответ. Реактивные окислители могут инактивировать хемотаксические факторы и служить для прекращения процесса привлечения нейтрофилов, ослабляя воспалительный процесс.
Высвобождение активных форм кислорода, белков гранул и цитокинов также может повредить местные ткани, что проявляется классическими признаками воспаления или приводит к необратимым нарушениям целостности и функций тканей.