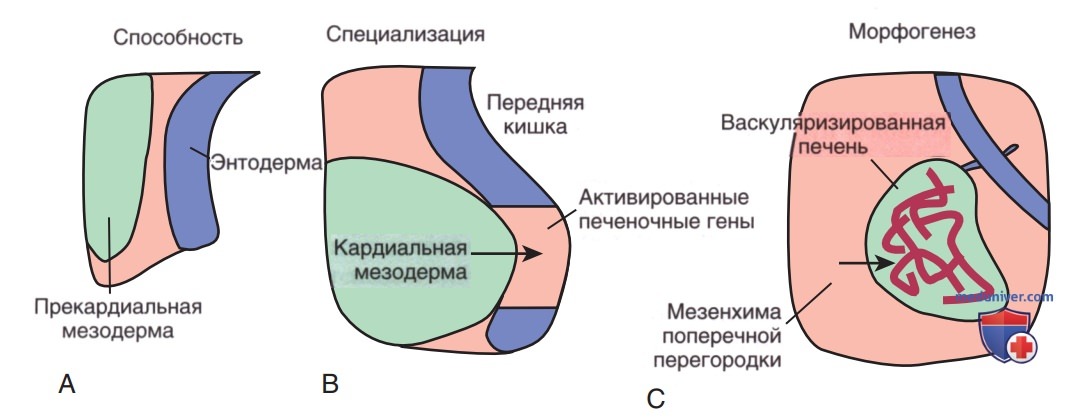
На ранних этапах эмбрионального развития в процессе гаструляции формируются три зародышевых листка (энтодерма, мезодерма и эктодерма). Печень и желчная система развиваются из эндодермальных клеток передней кишки, и это развитие можно разделить на три отдельных процесса (рис. 1). Во-первых, с помощью не вполне выясненных механизмов энтодерма передней кишки приобретает способность воспринимать сигналы, исходящие от кардиальной мезодермы.
Рисунок 1. Процессы, происходящие в раннем периоде развития печени: А — энтодерма передней кишки приобретает способность воспринимать сигналы, исходящие от кардиальной мезодермы; В — специфические клетки энтодермы передней кишки проходят через процесс специализации и активации печеночно-специфических генов под влиянием сигналов от мезодермы; С — морфогенез печени запускается по мере того, как прошедшие специализацию клетки мигрируют в поперечную перегородку под влиянием сигнальных молекул и экстрацеллюлярного матрикса, высвобождаемого клетками мезенхимы поперечной перегородки и примитивными эндотелиальными клетками.
Поступление этих мезодермальных сигналов в форме разл. факторов роста фибробластов и костных морфогенетических белков приводит к специализации клеток, потенциально способных к формированию ткани печени и активации генов, специфичных для нее. Во время этого решающего для развития печени периода «пионерные» факторы транскрипции, включая Foxa и Gata4, соединяются со специфичными сайтами связывания в компактном хроматине, открывают локальную структуру хроматина и маркируют гены как компетентные. Однако экспрессия этих генов произойдет только в случае, если они правильно индуцируется дополнительными факторами транскрипции.
Затем на 4-й неделе беременности эти совсем недавно ставшие специализированными клетки отслаиваются от энтодермы передней кишки и мигрируют в краниально-вентральном направлении в поперечную перегородку, инициируя морфогенез печени.
Рост и развитие новообразованной печени требует взаимодействия с эндотелиальными клетками. Важность некоторых белков для развития печени была продемонстрирована на животных моделях (табл. 1). Помимо этих белков, функциональную роль в регуляции экспрессии генов и в развитии гепатобилиарной системы на моделях рыбок данио-рерио и мышей играют микро-РНК, состоящие из небольших некодирующих одноцепочечных РНК.
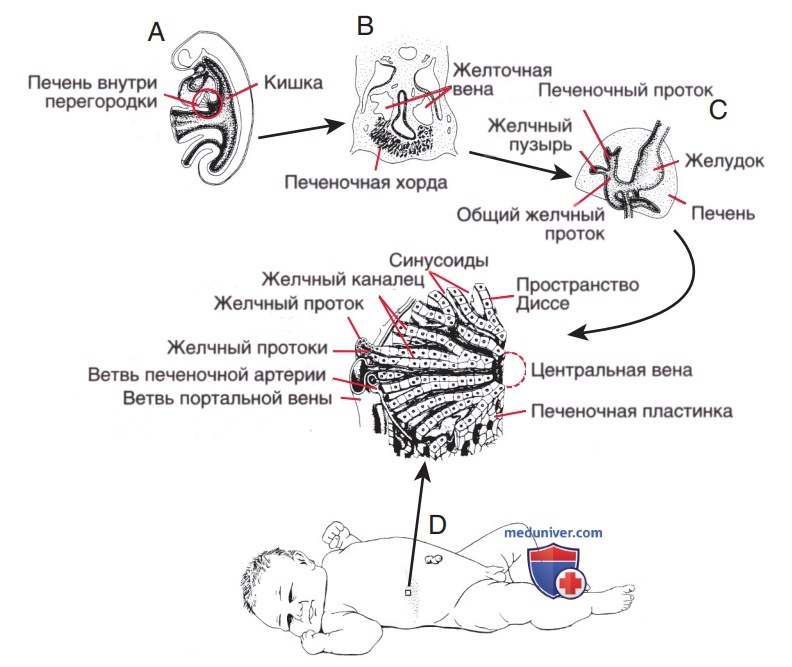
В результате пролиферации мигрирующих клеток внутри вентральной брыжейки формируются анастомозирующие печеночные тяжи, при этом сеть, состоящая из примитивных клеток печени, синусоидов и септальной мезенхимы формирует основной архитектурный паттерн дольки печени (рис. 2). Плотная краниальная часть печеночного дивертикула (pars hepatis) в конечном итоге формирует паренхиму печени и в/печеночные желчные протоки. Печеночные дольки можно различить уже на 6-й неделе беременности. Канальцевые структуры ЖВП, включая микроворсинки и комплексы межклеточных контактов, представляют собой специализированные участки мембран печеночных клеток.
Рисунок 2. Морфогенез печени: А — вентральный отросток дивертикула печени, выделившийся из энтодермы передней кишки эмбриона на сроке 3,5 нед.; В — между двумя желточными венами от растущего печеночного дивертикула отделяются эпителиальные (печеночные) тяжи, которые впоследствии станут паренхимой печени, вокруг них располагается эндотелий капилляров (синусоиды) (4 нед эмбрионального развития); С — гемисекция эмбриона на сроке 7,5 нед.; D — трехмерное изображение печеночной дольки новорожденного.
Они развиваются на очень ранних сроках беременности, и крупные канальцы, ограничиваемые несколькими гепатоцитами, различимы уже к 6-7-й неделе гестации.
Гепатоциты и клетки желчных протоков (холангиоциты) развиваются из одних и тех же кл.-предшественниц, гепатобластов. Сигнальный путь Notch, нарушенный при синдроме Алажилля (Alagille), стимулирует дифференцировку гепатобластов в клетки билиарного эпителия, в то время как фактор роста гепатоцитов препятствует этому процессу. Развитие в/печеночных желчных протоков определяется характером развития и ветвления воротной вены. Ок. 8-й недели беременности, начиная с ворот печени, примитивные гепатобласты, примыкающие к мезенхимальным клеткам, окружающим ветви воротной вены, образуют цилиндрический рукав, называемый протоковой пластинкой.
Начиная с 12-й недели беременности происходит ремоделирование протоковой пластинки, в ходе которого некоторые ее сегменты подвергаются тубулярному расширению, а избыточные клетки постепенно исчезают. Ветвление билиарного дерева продолжается на протяжении всего фетального периода, и к моменту рождения человека наиболее периферически расположенные ветви воротных вен все еще окружаются протоковыми пластинками. Требуется еще порядка 4 нед для того, чтобы произошла их трансформация в полноценные портальные протоки.
Нарушение ремоделирования протоковой пластинки приводит к сохранению ее примитивных конфигураций. Эта аномалия называется мальформацией протоковой пластинки.
Такие гистопатологические нарушения обнаруживаются в биоптатах печени при разл. заболеваниях, включая врожденный фиброз печени, болезнь Кароли (Caroli) и атрезию ЖВП.
Из каудальной части (pars cystica) печеночного дивертикула развиваются желчный пузырь, пузырный проток и общий желчный проток. Дистальные отделы правого и левого печеночных протоков развиваются из внепеченочных протоков, а проксимальные — из первых в/печеночных протоковых пластинок. Внепеченочные желчные протоки, а также находящееся в процессе развития в/печеночное билиарное дерево обеспечивают непрерывность и проходимость просвета уже на ранних этапах органогенеза (см. рис. 2).
Кровоток в печени плода обеспечивается печеночной артерией, а также воротной и пупочной венами, образующими портальный синус. Венозная кровь от воротной вены направляется преимущественно к правой доле печени, а от пупочной — в первую очередь влево. Венозный проток выполняет функцию шунта, отводящего кровь из воротной и пупочной вен в печеночную, минуя синусоидальную сеть. После рождения, в момент начала кормления, венозный проток облитерируется. У плода SaO2 в воротной вене ниже, чем в пуповинной венозной крови, соответственно, для правой доли печени характерна более низкая оксигенация и большая кроветворная активность, чем для левой.
Транспортная и метаболическая функция печени обеспечивается особой структурой печеночных тяжей, образуемых рядами гепатоцитов, разделенных синусоидами, и сходящихся к притокам печеночной вены (центральной вены), которая располагается в центре дольки (см. рис. 2). Эта структура обуславливает пути и закономерности притока разл. в-в в печень и их оттока из нее. Помимо поступления артериальной крови из системного кровотока, для печени также характерен венозный приток из ЖКТ через систему портальной вены. В свою очередь продукты гепатобилиарной системы выделяются двумя разл. способами: через печеночную вену, и через ЖВП обратно в кишечник.
Печенью секретируются плазменные белки и прочие компоненты плазмы. Абсорбированные и циркулирующие питательные в-ва поступают в печень через воротную вену/печеночную артерию, проходят через синусоиды и, минуя гепатоциты, попадают в системный кровоток из центральной вены. Компоненты желчи проходят через целый ряд расширяющихся каналов, попадая из желчных канальцев через желчные протоки в общий желчный проток.
Секрецию желчи впервые можно зафиксировать на 12-й неделе беременности. Состав желчи зависит от стадии развития. На ранних сроках в ней отмечается относительно низкое содержание ХС и фосфолипидов. Низкие концентрации желчных кислот, отсутствие желчных кислот бактериального происхождения (вторичных), а также присутствие некоторых особых желчных кислот отражают низкую скорость оттока желчи и незрелость путей синтеза желчных кислот.
Печень достигает максимального относительного размера, который составляет 10% от МТ плода, на 9-й неделе гестации. На ранних этапах эмбрионального развития печень — главное звено в процессе кроветворения. На 7-й неделе количество гемопоэтических клеток в зачатке печени превышает число функционирующих гепатоцитов. Эти незрелые гепатоциты имеют меньшие размеры, чем зрелые (20 мкм против 30-35 мкм), и содержат меньшее количество гликогена. Спустя недолгое время, по мере нарастания массы гепатоцитов и содержания в них гликогена, они начинают заметно преобладать в ткани органа.
У доношенных детей ко 2-му месяцу жизни кроветворение в печени уже практически отсутствует. Поскольку плотность гепатоцитов возрастает вместе со сроком беременности, удельный вес синусоидальной сети снижается. Печень составляет 5% МТР и лишь 2% у взрослого человека. У здоровых новорожденных некоторые метаболические процессы остаются незрелыми, отчасти из-за паттернов активности разл. ферментативных процессов у плода. Множество функций печени плода выполняются печенью матери, которая обеспечивает плод питательными в-вами, осуществляет выведение конечных продуктов метаболизма и токсинов. В печени плода главным образом происходят процессы выработки белков, необходимых для роста.
В дальнейшем основными функциями печени плода станут производство и хранение необходимых питательных в-в, экскреция желчи, налаживание процессов утилизации. В процессе внематочной адаптации ферменты необходимо синтезировать de novo. Регуляция этих процессов зависит от субстрата, гормонов, поступающих через плаценту, диеты и гормонального статуса в постнатальный период.
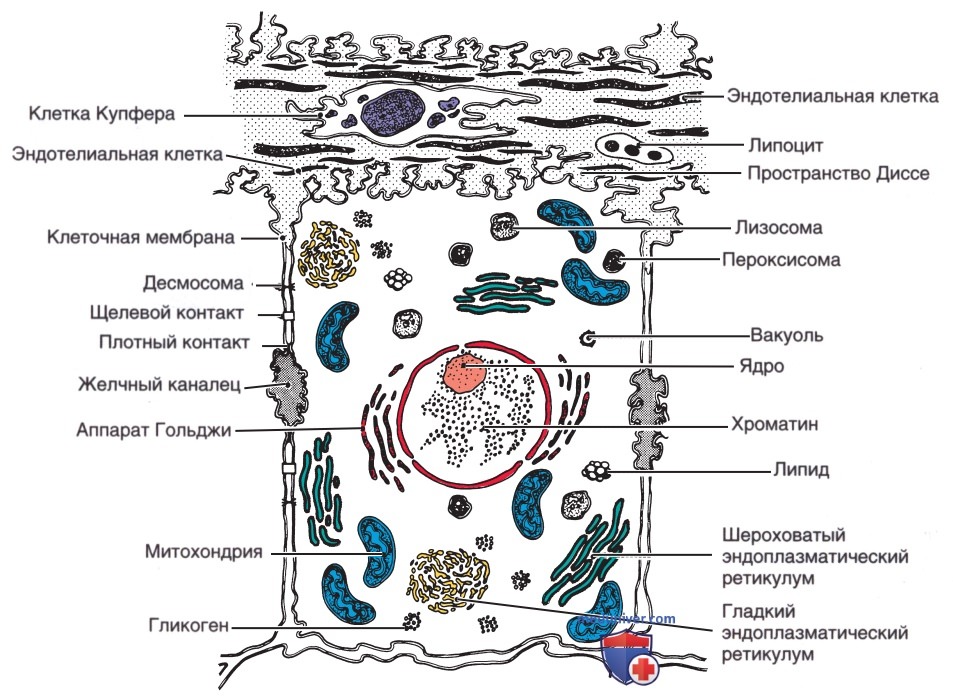
а) Ультраструктура печени. Гепатоциты обладают рядом ультраструктурных особенностей, которые отражают их биологические функции (рис. 3). Гепатоциты, как и др. эпителиальные клетки, поляризованы, что означает, что их структура и функция ориентированы в определенном направлении. Эта поляризация выражается, в частности, в том, что разл. области цитоплазматической мембраны гепатоцитов обладают специализированными функциями. Двунаправленный транспорт начинается на синусоидальной поверхности, куда попадают материалы, поступающие в печень через портальную систему, а в-ва, выделяемые печенью, покидают гепатоцит. Канальцевые мембраны соседствующих гепатоцитов образуют желчные канальцы, которые отграничиваются плотными контактами, предотвращающими транспорт секретируемых соединений обратно в синусоиду.
Рисунок 3. Схематическое изображение ультраструктуры и органелл гепатоцитов.
В свою очередь внутри гепатоцитов метаболическая и синтетическая активность реализуется разл. клеточными органеллами. Окисление и метаболизм гетерогенных классов субстратов, окисление жирных кислот, ключевые процессы глюконеогенеза, накопление и высвобождение энергии происходят в митохондриях, которые в большом количестве находятся в гепатоцитах.
Эндоплазматический ретикулум — непрерывная сеть, состоящая из канальцев с шероховатой и гладкой поверхностью, а также цистерн — место протекания множества процессов, включая синтез белков и триглицеридов, метаболизм ЛП. У плода отмечается низкая активность ферментов эндоплазматического ретикулума, что объясняет низкую эффективность метаболизма ксенобиотиков (ЛП). Аппарат Гольджи участвует в упаковке белков и, возможно, в секреции желчи. Пероксисомы гепатоцитов — одномембраннные цитоплазматические органеллы, содержащие такие ферменты, как оксидазы и каталазы, и ферменты, участвующие в метаболизме липидов и желчных кислот. Лизосомы содержат множество гидролаз, играющих важную роль во в/клеточном пищеварении.
Цитоскелет гепатоцитов, состоящий из актина и др. филаментов, распределен по клетке и сконцентрирован около плазматической мембраны. Микрофиламенты и микротрубочки играют важную роль в рецептор-опосредованном эндоцитозе, секреции желчи, поддержании архитектуры и подвижности гепатоцитов.
б) Метаболические функции печени:
1. Метаболизм углеводов. Печень регулирует уровень глюкозы в сыворотке крови посредством нескольких процессов, включая хранение ее излишков в виде гликогена, полимера глюкозы, быстро гидролизуемого обратно во время голодания. Для поддержания уровня глюкозы в сыворотке гепатоциты продуцируют свободную глюкозу путем гликогеноли-за/глюконеогенеза. Сразу после рождения ребенок зависим от процесса гликогенолиза в печени. В печени плода активность глюконеогенеза низка, однако она быстро нарастает после рождения. Синтез гликогена у плода начинается на ~9-й неделе беременности, и именно на этом сроке он аккумулируется быстрее всего, и спустя совсем немного времени в печени плода содержится в 2-3 раза больше гликогена, чем в печени взрослого человека.
Большая часть запасенного гликогена будет использована в раннем постнатальном периоде. Повторное накопление гликогена начинается на ~2-й неделе постнатального периода, и его запасы у здоровых доношенных детей достигают уровня взрослого на ~3-й неделе. У недоношенных детей уровни глюкозы в сыворотке могут колебаться по той причине, что эффективная регуляция процессов синтеза, хранения и утилизации гликогена развивается только ближе к концу доношенной беременности. Алиментарные углеводы, такие как галактоза, также превращаются в глюкозу, однако уровень глюкозы в раннем возрасте сильно зависит от глюконеогенеза, особенно в условиях ограниченных запасов гликогена.
2. Метаболизм белков. В период быстрого роста плода специфические декарбоксилазы, ограничивающие скорость биосинтеза физиол. важных полиаминов, обладают более высокой активностью, чем в печени взрослого человека. Скорость синтеза альбумина и секреторных белков в развивающейся печени параллельна количественным изменениям в эндоплазматическом ретикулуме. Синтез альбумина у плода человека начинается на 7-8-й неделе и увеличивается обратно пропорционально синтезу АФП, главного фетального белка. К 3-4 мес беременности печень плода способна продуцировать фибриноген, трансферрин и ЛПНП. С этого момента в плазме плода содержатся все основные классы белков, однако в значительно меньших, чем у взрослых людей, концентрациях.
Постнатальные паттерны синтеза белков различаются в зависимости от класса. Уровни липопротеинов всех классов резко повышаются уже в течение первой недели после рождения и достигают уровней, которые почти не будут меняться вплоть до периода полового созревания. У новорожденных отмечается низкое содержание альбумина (~2,5 г/дл), оно достигает уровня взрослого человека (~3,5 г/дл) через несколько месяцев. Уровни церулоплазмина и факторов комплемента медленно повышаются до уровней взрослого в течение первого года жизни. Уровень трансферрина, напротив, при рождении аналогичен таковому у взрослого человека, затем в течение 3-5 мес он снижается, а далее вновь повышается до достижения конечной концентрации. Низкий уровень активности специфических белков влияет на питание младенца.
К примеру, низкая активность цистатионин-γ-лиазы (цистатионазы) негативно влияет на процесс транссульфирования, посредством которого поступающий с пищей метионин превращается в цистеин. Следовательно, будет необходимо поступление цистеина с пищей. Такая же ситуация может сложиться и в отношении др. серосодержащих аминокислот, напр. таурина.
3. Метаболизм липидов. Окисление жирных кислот, вместе с гликогенолизом и глюконеогенезом, является основным источником энергии на раннем этапе жизни. Новорожденные относительно плохо переносят длительное голодание, отчасти из-за ограниченной способности их печени к кетогенезу. Способность печени окислять жирные кислоты формируется быстро, в первые несколько дней жизни. Молоко — основной источник калорий в раннем возрасте, и именно вследствие высокого содержания жиров и низкого содержания углеводов в рационе на данном этапе жизни для поддержания уровня глюкозы в крови необходим активный глюконеогенез.
В условиях ограниченного поступления глюкозы продукция кетоновых тел из эндогенных жирных кислот может обеспечить энергию, необходимую для глюконеогенеза в печени, стать альтернативным источником энергии для метаболизма ГМ. При наличии лишних углеводов печень продуцирует триглицериды. Метаболические процессы с участием липидов и липопротеинов происходят преимущественно в печени, поэтому незрелость/заболевания печени оказывают существенное влияние на их концентрацию.
4. Биотрансформация. У новорожденных снижена способность к метаболизму и обезвреживанию определенных ЛП по причине слабого развития микросомального компонента печени, который представляет собой место протекания специфических окислительных, восстановительных, гидролитических реакций, а также реакций конъюгации, необходимых для вышеуказанных биотрансформаций. Основные компоненты монооксигеназной системы, такие как цитохром Р450, цитохром-с-редуктаза и восстановленная форма никотинамидадениндинуклеотидфосфата, в микросомах плода присутствуют в низких концентрациях. У доношенных детей печеночная уридиндифосфатглюкуронозилтрансфераза и ферменты, участвующие в окислении полициклических ароматических углеводородов, характеризуются очень низким уровнем экспрессии.
Возрастные различия в фармакокинетике варьируют от соединения к соединению. Так, период полувыведения парацетамола у новорожденного аналогичен таковому у взрослого человека, тогда как, напр., период полувыведения теофиллина у недоношенного ребенка составляет ~100 ч по сравнению с 5-6 ч у взрослого. Именно эти различия в метаболизме, а также такие факторы, как связывание с белками плазмы и почечный клиренс, и определяют дозировку ЛП, которая позволит добиться максимального эффекта и избежать токсичности. Яркими примерами предрасположенности новорожденных к лекарственной токсичности являются реакции на хлорамфеникол (синдром «серого ребенка»), бензоиловый спирт и его метаболиты. Эти реакции обусловлены неэффективной конъюгацией глюкуронида и глицина соответственно.
Низкие концентрации антиоксидантов (витамин Е, супероксиддисмутаза, глутатионпероксидаза) в печени плода и новорожденного приводят к повышенной восприимчивости к пагубным проявлениям токсичности кислорода и окислительному повреждению вследствие перекисного окисления липидов.
Реакции конъюгации, превращающие ЛП/их метаболиты в водорастворимые формы, которые могут выводиться с желчью, также катализируются микросомальными ферментами печени. У новорожденных детей наблюдается сниженная активность печеночной уридиндифосфатглюкуронозилтрансферазы, которая превращает неконъюгированный билирубин в легко выводимый конъюгат глюкуронида, а также является ферментом, ограничивающим скорость выведения билирубина. Трансфераза в постнатальном периоде быстро становится активной, независимо от доношенности ребенка, что позволяет сделать предположение о том, что главную роль в постнатальном развитии активности этого фермента играют факторы, связанные с рождением, а не возрастом плода. Микросомальную активность можно стимулировать введением фенобарбитала, рифампицина и др. индукторов цитохрома Р450. И, напротив, такие ЛП, как циметидин, могут ингибировать микросомальную активность Р450.
5. Экскреторная функция печени. Выделительная функция печени и отток желчи тесно взаимосвязаны с печеночной экскрецией желчных кислот и кишечно-печеночной рециркуляцией. Желчные кислоты, основные продукты деградации ХС, включаются в смешанные мицеллы вместе с ХС и фосфолипидами. Эти мицеллы являются эффективным носителем, необходимым для солюбилизации и всасывания липофильных соединений, таких как пищевые жиры и жирорастворимые витамины, в кишечнике. Секреция желчных кислот клетками печени — основной фактор, определяющий отток желчи у взрослых животных. Соответственно, зрелость процессов обмена желчных кислот влияет и на общую выделительную функцию печени, включая выведение эндогенных и экзогенных соединений с желчью.
В печени человека синтезируются две основные желчные кислоты, холевая и хенодезоксихолевая. Перед выведением из организма они связываются с глицином и таурином. В ответ на прием пищи происходит сокращение желчного пузыря, за счет которого желчные кислоты выбрасываются в кишечник, где способствуют перевариванию и всасыванию жиров. Приняв участие в опосредованном переваривании жиров, желчные кислоты реабсорбируются из терминального отдела подвздошной кишки при помощи специфических процессов активного транспорта. Они вновь попадают в печень через воротную вену, поглощаются клетками печени и повторно включаются в состав желчи. У взрослого человека 90-95% пула циркулирующих желчных кислот включены в процесс кишечнопеченочной циркуляции.
Желчные кислоты, нереабсорбировавшиеся в подвздошной кишке, достигают толстой кишки, где бактериальная флора путем дигидроксилирования и деконъюгации продуцирует вторичные желчные кислоты, дезоксихолат и литохолат. Т.о., в состав желчи взрослого человека входят как первичные, так и вторичные желчные кислоты, реабсорбируемые в дистальных отделах ЖКТ.
В/просветные концентрации желчных кислот у детей непосредственно после рождения низкие, однако их уровни довольно быстро повышаются. Расширение пула желчных кислот играет очень важную роль, поскольку они абсолютно необходимы для стимуляции оттока желчи и абсорбции липидов, главной составляющей пищи новорожденного. Такие ядерные рецепторы, как рецептор фарнезоида X, контролируют в/печеночный гомеостаз желчных кислот посредством нескольких механизмов, включая регуляцию экспрессии генов, кодирующих два ключевых белка: холестерин-7α-гидроксилазу (CYP7A1) и насос выведения солей желчных кислот (BSEP). Эти белки играют важную роль в синтезе желчных кислот и канальцевой секреции соответственно. Неонатальная экспрессия этих ядерных рецепторов варьирует в зависимости от изучаемой животной модели, а у человека остается сравнительно малоизученной.
По причине недостаточной реабсорбции желчных кислот в подвздошной кишке, а также низкой скорости их выведения из портальной крови концентрации желчных кислот в сыворотке здоровых новорожденных обычно бывают повышенными, зачастую настолько, что у пожилых людей они могли бы указывать на наличие заболевания печени. У новорожденных с низкой МТР, а также у доношенных детей после перенесенного в перинатальном периоде стресса, напр. гипоксии/инфекции, часто встречаются физиол. транзиторный холестаз и стеаторея, которые редко наблюдаются у здоровых доношенных новорожденных.
Множество процессов, обусловленных незрелостью новорожденного в плане морфогенеза и функции печени, о чем уже говорилось ранее, влияет на повышенную восприимчивость этих детей к заболеваниям печени, вызванных парентеральным питанием. Уменьшенный пул солей желчных кислот, истощение печеночного глутатиона и недостаточная сульфатация способствуют выработке токсичной литохолевой желчной кислоты, развитию холестаза, и в то же время дефицит незаменимых аминокислот, таких как таурин и цистеин, и избыточная инфузия липидов у таких младенцев могут привести к стеатозу печени. После окончания неонатального периода нарушения обмена желчных кислот могут оказывать разл. влияние на гепатобилиарную и кишечную функции (табл. 2).
Видео эмбриогенез, строение, гистология печени и гепатоцитов