Методы хромосомного анализа (цитогенетики) - кратко с точки зрения педиатрии
Клиническая цитогенетика занимается изучением хромосом: их структуры, функции, наследования и аномалий. Хромосомные аномалии распространены достаточно широко и обнаруживаются у 1-2% живорожденных, 5% мертворожденных младенцев и в 50% случаев ранней потери плода в I триместре беременности (табл. 1). Хромосомные аномалии чаще встречаются среди людей с умственной отсталостью и играют значительную роль в развитии некоторых новообразований.
Хромосомный анализ показан лицам с множественными ВПР, клинически анормальным фенотипом / дисморфизмом / умственной отсталостью. Отдельными показаниями для проведения исследования являются: возраст матери >35 лет, множественные отклонения от нормы при УЗИ плода (пренатальное обследование), множественные врожденные аномалии, необъяснимая задержка роста плода, проблемы в росте и развитии после рождения, неопределенные гениталии, необъяснимая умственная отсталость с/без сопутствующих анатомических аномалий, первичная аменорея / бесплодие, повторяющиеся выкидыши (≥3) / наличие в анамнезе мертворождений / неонатальных смертей, родственник первой степени родства с известной/предполагаемой структурной хромосомной аномалией, клинические данные, соответствующие известной патологии, некоторые ЗНО и синдромы хромосомной нестабильности (напр., синдром Блума, анемия Фанкони).
Цитогенетические исследования обычно проводят на лимфоцитах периферической крови, хотя можно также использовать культивированные фибробласты, полученные при биопсии кожи. Пренатальные (фетальные) исследования хромосом выполняются с использованием клеток, полученных из околоплодных вод (амниоцитов), ткани ворсинок хориона и крови плода / в случае доимплантационной диагностики путем биопсии бластомера (на стадии дробления эмбриона), биопсии полярного тела / биопсии бластоцисты. Цитогенетические исследования костного мозга играют важную роль в наблюдении за опухолями, особенно у пациентов с лейкозами. Они применяются для определения индукции ремиссии, эффективности терапии / в отдельных случаях возникновения рецидивов.
Хромосомные аномалии возникают вследствие ошибок во время деления клеток и подразделяются на числовые и структурные. Существует два типа деления клеток: митоз, который происходит в большинстве соматических клеток, и мейоз, который ограничивается половыми клетками. В процессе митоза из одной родительской клетки образуются две генетически идентичные дочерние клетки. Дублирование ДНК уже произошло во время интерфазы в S-фазе клеточного цикла (синтез ДНК). Следовательно, в начале митоза хромосомы состоят из двух двойных цепей ДНК, соединенных в области центромеры, известных как сестринские хроматиды. Митоз можно разделить на четыре стадии: профаза, метафаза, анафаза и телофаза. Профаза характеризуется началом конденсации ДНК. Во время профазы ядерная оболочка и ядрышко исчезают, и формируется веретено деления.
Во время метафазы хромосомы максимально конденсированы и отчетливо видны как отдельные структуры. Хромосомы выстраиваются в центре клетки, а нити веретена соединяются с центромерами каждой хромосомы и расходятся к центриолям, образуя фигуру митоза. Во время анафазы хромосомы делятся вдоль своих продольных осей с образованием двух дочерних хроматид, которые затем мигрируют к противоположным полюсам клетки. Телофаза характеризуется образованием двух новых ядерных оболочек и ядрышек, дупликацией центриолей и расщеплением цитоплазмы с образованием двух дочерних клеток.
Мейоз начинается в женском ооците во время жизни плода и завершается спустя годы/десятилетия. У мужчин он начинается в определенной сперматогониальной клетке где-то между подростковым возрастом и взрослой жизнью и завершается за несколько дней. Мейозу предшествует репликация ДНК, поэтому вначале каждая из 46 хромосом состоит из двух хроматид. Во время мейоза диплоидная клетка (2n = 46 хромосом) делится с образованием четырех гаплоидных клеток (n = 23 хромосомы). Мейоз состоит из двух основных фаз деления клеток. Во время мейоза I каждая из гомологичных хромосом спаривается т.о., чтобы обеспечить генетическую рекомбинацию, т.е. обмен между двумя цепями ДНК (кроссинговер).
В результате происходит перегруппировка генетической информации для рекомбинированных хромосом и обеспечивается дальнейшее генетическое разнообразие. Затем каждая дочерняя клетка получает по 1 из 23 гомологичных хромосом. В оогенезе одна из дочерних клеток получает большую часть цитоплазмы и трансформируется в яйцеклетку, тогда как другая, меньшая клетка становится первым полярным тельцем. Мейоз II подобен митотическому делению, но без предшествующей фазы дупликации (репликации) ДНК. Каждая из 23 хромосом делится продольно, а гомологичные хроматиды мигрируют к противоположным полюсам клетки. В результате образуется четыре сперматогония (у мужчин) / яйцеклетка и два полярных тельца (у женщин), каждый с гаплоидным (n = 23) набором хромосом. Следовательно, мейоз реализует две важные функции: осуществляет редукцию числа хромосом с диплоидного (46) до гаплоидного (23), так что при оплодотворении диплоидное число восстанавливается и обеспечивает генетическую рекомбинацию.
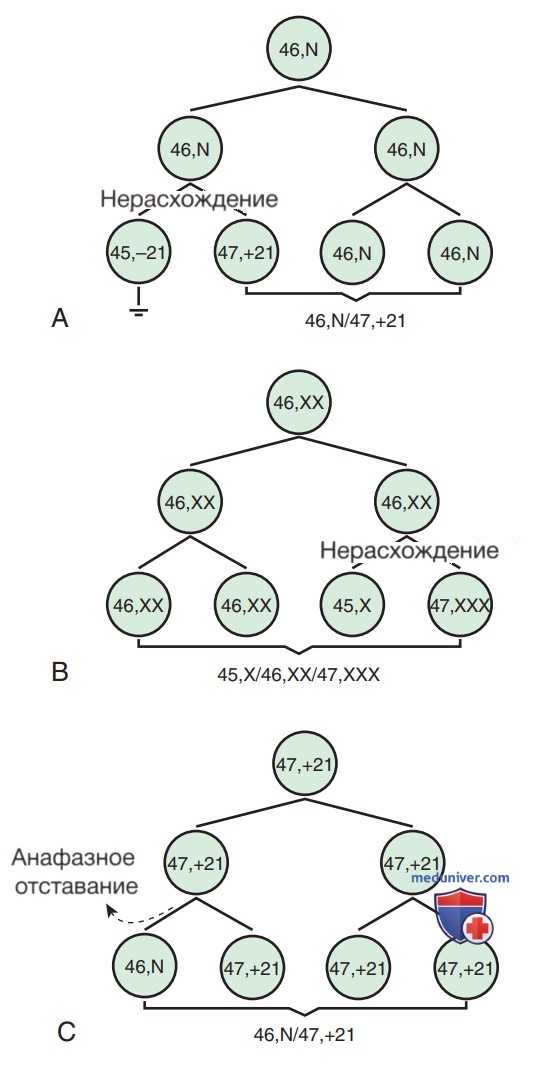
Во время мейоза/митоза могут происходить две наиболее частые ошибки деления клеток, которые могут привести к нарушению числа хромосом. Первая ошибка — нерасхождение хромосом, когда две хромосомы не разъединяются и вместе мигрируют в одну из новых клеток, в результате образуется одна клетка с двумя копиями хромосом и вторая без копии. Второй ошибкой является анафазное отставание, при котором хроматида/хромосома из-за своей неспособности двигаться достаточно быстро во время анафазы утрачивается и не включается в одну из новых дочерних клеток (рис. 1).
Рисунок 1. Образование мозаицизма: А — постзиготическое нерасхождение у изначально нормального продукта зачатия. В этом примере одна клеточная линия (моносомная 21) впоследствии утеряна с образованием окончательного кариотипа 46,N/47,+21; В — постзиготное нерасхождение в первоначально 46,XX продукте зачатия, в результате возникает мозаицизм 45,Х/46,ХХ/47,ХХХ; C — постзиготная задержка анафазы у первоначально 47,+21 продукта зачатия.
Для хромосомного анализа клетки культивируют (в течение различных периодов времени в зависимости от типа клеток) со стимуляцией / без нее, а затем искусственно задерживают митоз во время метафазы/прометафазы, подвергают воздействию гипотонического р-ра, позволяющего разрушить оболочку ядерной клетки и обеспечить правильное распределение хромосом для анализа, проводят фиксацию, предобработку и окончательную окраску. Наиболее часто используемым методом дифференциального окрашивания хромосом является GTG-бэндинг (разновидность G-бэндинга, включает обработку препарата трипсином и окраску красителем Гимза), который дает уникальную комбинацию темных (G-«+») и светлых (G-«-») полос, позволяющую идентифицировать каждую из 23 пар анализируемых хромосом.
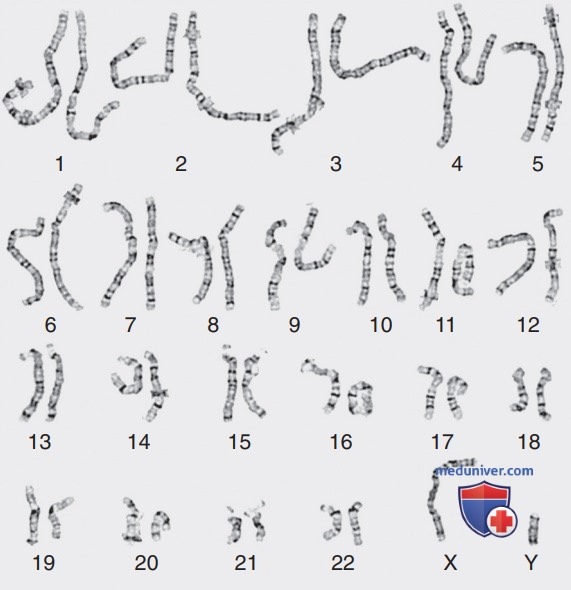
Распределение метафазных хромосом сначала рассматривается под микроскопом, а затем их изображения фо-тографируются/фиксируются видеокамерой и сохраняются на компьютере для последующего анализа. У человека имеется 46 хромосом / 23 пары, которые классифицируются как аутосомы для хромосом 1-22, и половые хромосомы, часто называемые половым комплементом: XX для женщин и XY для мужчин. Гомологичные хромосомы из метафазной пластинки раскладывают по парам и располагают в определенном порядке для сборки кариотипа в соответствии с четко определенными, стандартными правилами, установленными Международной системой цитогенетической номенклатуры человека (ISCN), при этом хромосома 1 является самой большой, а 22 — самой маленькой. По номенклатуре описание кариотипа включает общее количество хромосом, за которым следует состав половых хромосом. Нормальный кариотип — 46,XX для женщин и 46,XY для мужчин (рис. 2). Аномалии записывают после половых хромосом.
Рисунок 2. Кариотип здорового мужчины с разрешением 550-600 полос. Чем длиннее хромосомы, захваченные в метафазе / иногда в прометафазе, тем больше полос можно визуализировать
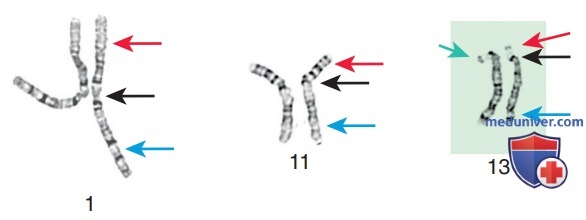
Хотя международная система номенклатуры хромосом человека в основном базируется на длине и исчерченности каждой хромосомы, положение центромеры относительно концов хромосомы также является отличительным признаком (рис. 3). Центромера делит хромосому на две части, причем короткое плечо обозначено как р-плечо, а длинное — как q-плечо. Знак ± перед номером хромосомы указывает соответственно на наличие лишней/отсут-ствующей хромосомы. В табл. 2 перечислены некоторые сокращения, используемые для описания хромосом и их аномалий.
Рисунок 3. Пример различных типов хромосом в зависимости от положения центромеры. Слева находятся хромосомы 1-й пары с центромерой, одинаково удаленной от короткого и длинного плеча (также известной как метацентрическая). В центре находятся хромосомы 11-й пары, которая является субметацентрической. Справа — 13-я пара хромосом, которая является примером акроцентрической хромосомы. У акроцентрических хромосом очень маленькое короткое плечо, спутничные нити и спутники, состоящие из сателлитной ДНК. Черная стрелка указывает положение центромеры. Синяя стрелка — длинное плечо хромосомы. Красная стрелка — короткое плечо хромосомы. Зеленой стрелкой выделены спутники, состоящие из повторов ДНК. Светлая область между коротким плечом и спутником называется спутничной нитью
Метафазные хромосомы обычно дают разрешение 450-550 полос. Профазные и прометафазные хромосомы длиннее, менее уплотнены и часто содержат 550-850 полос. Анализ с высоким разрешением может обнаруживать небольшие хромосомные аномалии, хотя в основном он был заменен методом молекулярного кариотипирования на ДНК-микроматрицах (метод CGH/aCGH).
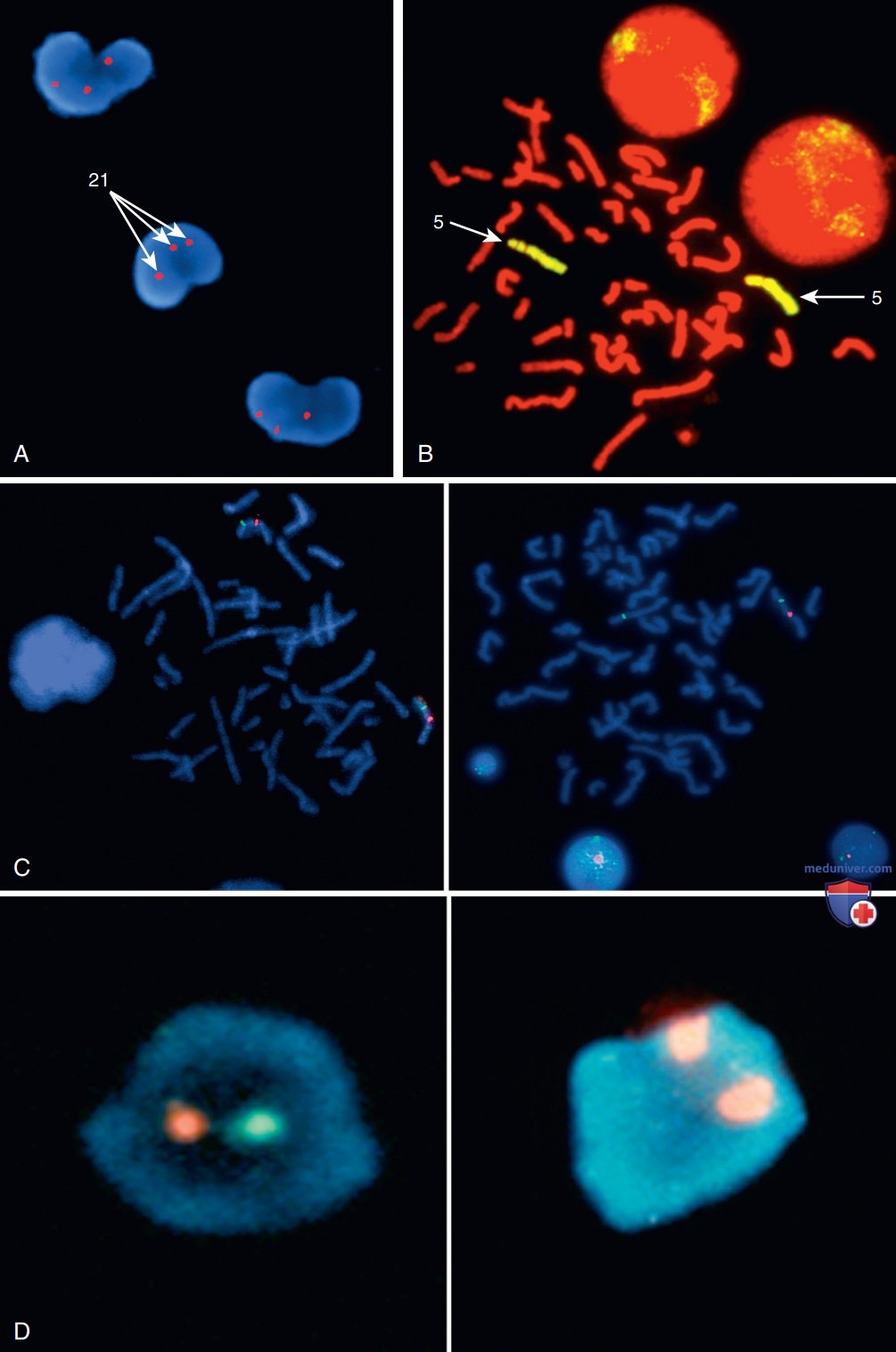
Молекулярные методы (FISH, СМА, aCGH) заполнили значительный пробел в диагностике криптических хромосомных аномалий. Эти методы способны выявлять малейшие отклонения, которые часто не определяются при стандартных цитогенетических исследованиях. Флуоресцентная гибридизация (FISH) in situ используется для идентификации наличия, отсутствия/перестрой-ки определенных сегментов ДНК и выполняется с помощью ДНК-зондов, специфичных для гена / участка гена. В клинических условиях используются несколько FISH-зондов: на уникальные последовательности / однокопийные зонды, зонды на повторяющиеся последовательности (α-сателлиты перицентромерных участков) и мультикопийные зонды (специфичные для отдельных участков хромосом / цельнохромосомные) (рис. 4).
Рисунок 4. А — FISH-анализ интерфазных клеток периферической крови пациента с синдромом Дауна с использованием специфичного зонда для хромосомы 21. Три красных сигнала обозначают наличие трех хромосом 21; В — FISH-анализ препарата метафазных хромосом от клинически здорового человека, с использованием полнохромосомного зонда, специфичного для хромосомы 5. Обе хромосомы 5 полностью окрашены (желтым цветом) по всей своей длине; C — FISH на метафазных клетках с использованием зонда к уникальной последовательности, который гибридизируется с геном эластина на хромосоме 7q11.23, в пределах критического участка генома, ответственного за развитие синдрома Вильямса. Эластиновый зонд помечен красным, а контрольный зонд на хромосоме 7 помечен зеленым. На рисунке слева изображена нормальная гибридизация с хромосомой 7, с двумя сигналами для области эластина и двумя для контрольного зонда. На рисунке справа: справа показана нормальная хромосома с сигналами контроля и эластина, а слева — делеция хромосомы 7, о чем свидетельствует единственный сигнал контрольного зонда. Это изображение принадлежит пациенту с делецией генетического материала из области, ответственной за развитие синдрома Вильямса; D — FISH в интерфазных клетках с использованием ДНК-зондов, которые гибридизуются с повторяющимися последовательностями α-сателлитов в перицентромерной области половых хромосом. Слева: интерфазные клетки с двумя сигналами, один из которых отмечен красным для Х-хромосомы и зеленым для Y-хромосомы, что соответствует нормальному набору мужских хромосом. Справа: интерфазная клетка, показывающая два красных сигнала для Х-хромосомы, соответствующие нормальному набору женских хромосом
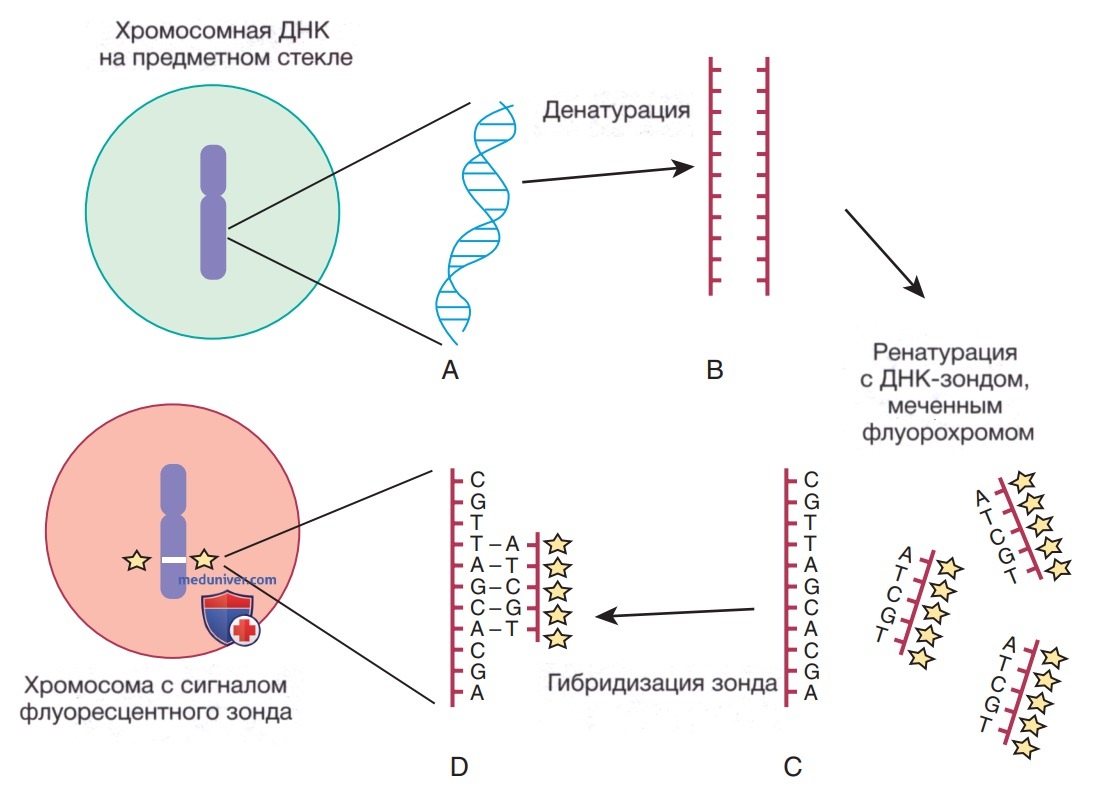
Метод FISH заключается в использовании уникальной известной последовательности ДНК/зонда, меченного флуоресцентным красителем, который комплементарен исследуемому на предмет болезни участку ДНК. Меченый зонд гибридизуется с ДНК на предметном стекле микроскопа, как правило, это метафазная/интерфазная хромосомная ДНК. В момент гибридизации зонда с его комплементарной последовательностью ДНК его можно визуализировать с помощью флуоресцентной микроскопии (рис. 5). При анализе метафазных хромосом можно задокументировать точное хромосомное положение каждой копии зонда, а также, довольно часто, количество копий (делеций и дупликаций) последовательностей ДНК. Когда исследуемые сегменты (как при дупликациях) расположены близко друг к другу, только интерфазные клетки могут точно определить наличие >2 копий/сигналов, поскольку в метафазных клетках некоторые дупликации могут ошибочно расцениваться как один сигнал.
Рисунок 5. Метод FISH включает денатурацию двухцепочечной ДНК, присутствующей в метафазных хромосомах / интерфазных ядрах цитогенетических препаратов на слайдах (А), в одноцепочечную ДНК (В). Затем фиксированную к предметному стеклу (in situ) ДНК ренатурируют / повторно отжигают в присутствии избыточных копий одноцепочечной, меченной флуорохромом последовательности пар оснований ДНК/зонда (С). Зонд отжигается/«гибридизируется» с участками комплементарной последовательности ДНК (D) в хромосомном геноме. Сигнал зонда визуализируется и отображается на хромосоме с помощью флуоресцентной микроскопии
Хромосомные перестройки, охватывающие <5 млн п.о. (5 Мб), не м.б. обнаружены обычными цитогенетическими методами. Первоначально метод FISH использовался для обнаружения малых делеций ДНК размером <50-200 Кб и облегчил раннее клиническое обнаружение ряда микроделеционных синдромов. Некоторые зонды FISH гибридизуются с повторяющимися последовательностями, расположенными в перицентромерных областях. Перицентромерные зонды до сих пор широко используются для быстрой идентификации некоторых трисомий в интерфазных клетках мазков крови / даже для быстрого анализа клеток плода, полученных с помощью амниоцентеза. Такие зонды существуют для хромосом 13, 18 и 21, а также для пары половых хромосом X и Y (см. рис. 4). Что касается выявления геномных нарушений, FISH больше не является тестом первой линии, но его роль важна для подтверждения результатов микрочипов.
Т.о., FISH предназначен для 1) подтверждающих исследований патологий, обнаруженных с помощью хромосомного микроматричного анализа; 2) быстрого пренатального скрининга на межфазные клетки околоплодных вод; 3) межфазного мазка крови для определения пола новорожденных с неоднозначными гениталиями.
Сравнительная геномная гибридизация на микроматрицах (aCGH) и хромосомный микроматричный анализ — молекулярные методы, заключающиеся в дифференцированном окрашивании ДНК пациента флуоресцентным красителем (зеленый флуорофор) и нормальной эталонной ДНК красным флуорофором (рис. 6). Олигонуклеотиды (короткие сегменты ДНК), охватывающие весь геном, наносятся на предметное стекло / сетку микрочипов. Равные количества двух образцов ДНК, помеченных флуоресцентными метками, смешивают и измеряют соотношение зеленой и красной флуоресценции вдоль каждой тестируемой области. Области амплификации ДНК пациента демонстрируют избыточную зеленую флуоресценцию, а области потери — избыточную красную флуоресценцию. Если ДНК пациента и контрольная ДНК представлены одинаково, соотношение зеленого и красного составляет 1:1, а тестируемые области отображаются желтым цветом (рис. 5).
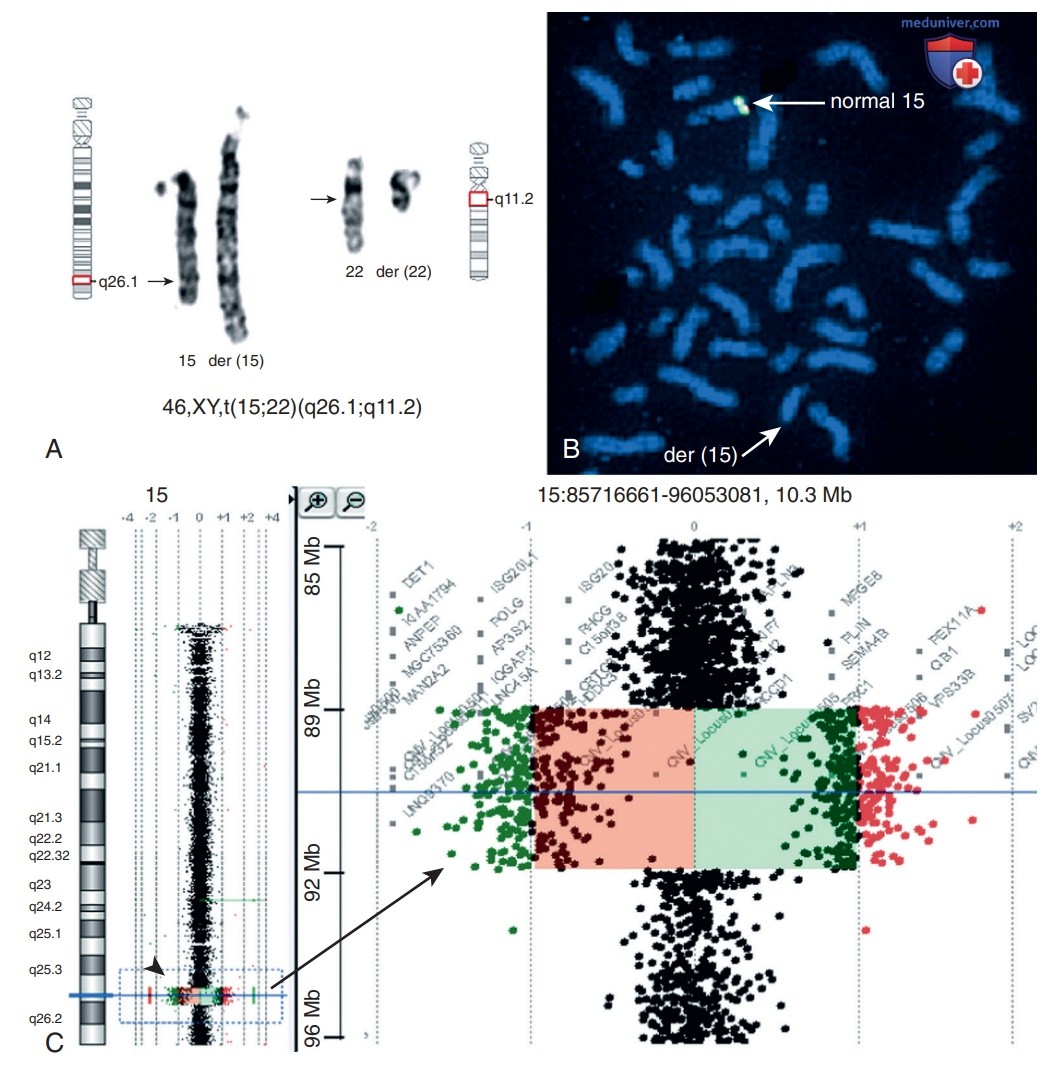
Рисунок 6. Пример скрытой микроделеции в точке разрыва транслокации при цитогенетически сбалансированной транслокации у пациента с задержкой развития и нарушением роста: A — частичный кариотип показывает t(15;22)(q26.1;q11.2); B — FISH с клонами 2019 (зеленый) и 354М14 (красный) в 15q26.1; стрелки указывают на сигналы, присутствующие только на нормальной хромосоме 15, что указывает на делецию на der(15); C — двухцветная матричная сравнительная геномная гибридизация с заменой красителя на олигозонды 244 К; стрелка указывает на делецию 3,3-Мб в хромосоме 15q26.1 -q26.2 (вид крупным планом)
В настоящее время возможно обнаружение на уровне разрешения одного экзона, в зависимости от используемых микроматриц. В ближайшем будущем определение числа копий может усовершенствоваться до такой степени, что будет обнаруживаться с помощью секвенирования следующего поколения в контексте полногеномного секвенирования.
К преимуществам метода aCGH относятся способность одновременно тестировать все критические, ответственные за развитие заболеваний участки генома, обнаруживать дупликации и делеции, которые в настоящее время не распознаются как области, вызывающие рецидивы заболевания в семье и определяемые методом FISH, а также обнаруживать синдромы делеции как одного, так и смежных генов. Кроме того, метод aCGH не всегда требует, чтобы культура клеток генерировала достаточное количество ДНК, что м.б. важно в контексте пренатального тестирования из-за ограничений во времени. Недостатком метода aCGH является то, что он не обнаруживает сбалансированные транслокации/инверсии и может не обнаружить низкие уровни хромосомного мозаицизма. Среди различных типов aCGH одни более таргетированы, чем другие.
Таргетированный метод aCGH м.б. эффективным для обнаружения известных клиницистам критических хромосомных аберраций, обычно ассоциируемых с известными фенотипами заболеваний. Полногеномные матрицы нацелены на весь геном и позволяют лучше и плотнее покрывать равномерно распределенные области генома.
Их недостатком является то, что интерпретация деле-ций/дупликаций м.б. затруднена, если они затрагивают области, о которых ранее не было известно, что они вовлечены в патологический процесс. Часто используемые в клинических условиях матрицы — однонуклеотидные полиморфизмы (single nucleotide polymorphism — SNP). SNP представляют собой полиморфные вариации между двумя нуклеотидами, и при массовом параллельном анализе они могут предоставить ценную клиническую информацию. В геноме человека обычно обнаруживаются несколько миллионов SNP.
Матрицы SNP могут помочь в обнаружении однородительских дисомий (т.е. генетическая информация, полученная только от одного родителя), а также кровное родство в семье. Многие массивы, в настоящее время используемые в клинической практике, сочетают в себе использование олигонуклеотидов для обнаружения вариаций числа копий в сочетании с SNP. Существует множество вариаций числа копий, способных вызвать делецию/дупликацию в геноме человека. Т.о., большинство обнаруженных генетических аномалий, если они не связаны с хорошо известными клиническими фенотипами, требуют обследования родителей, потому что обнаруженная унаследованная вариация числа копий м.б. доброкачественной / случайным полиморфным вариантом. Мутация de novo (т.е. обнаруживаемая только у ребенка, а не у родителей) часто бывает более значительной, если она связана с аномальным фенотипом, обнаруживаемым только у ребенка, а также если она включает гены с важными функциями.
aCGH — очень ценная технология сама по себе / в сочетании с FISH и обычными хромосомными исследованиями (рис. 7).
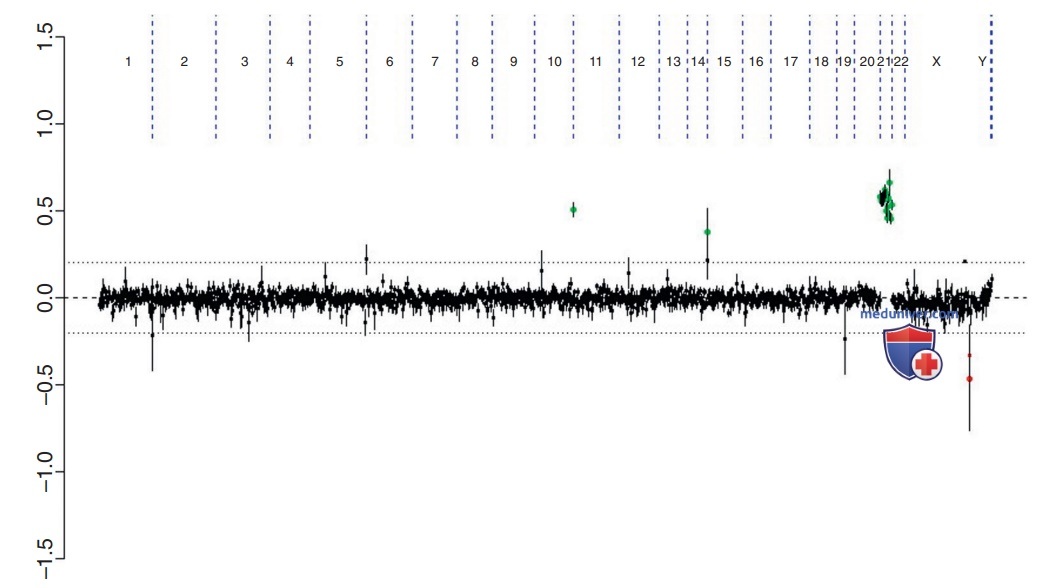
Рисунок 7. aCGH у пациентки с синдромом Дауна. Каждая черная точка представляет собой часть сегмента ДНК, специфичную для разного расположения хромосомы. Считается, что большинство точек, отображаемых между осями 0,0 и 0,2, находятся в пределах нормы. Исключения часто являются результатом полиморфных вариаций. Группа зеленых точек в кластерах на хромосоме 21 и выше 0,5. Они представляют собой увеличение числа копий сегментов ДНК для хромосомы 21, наблюдаемое при синдроме Дауна, соответствующее трисомии 21