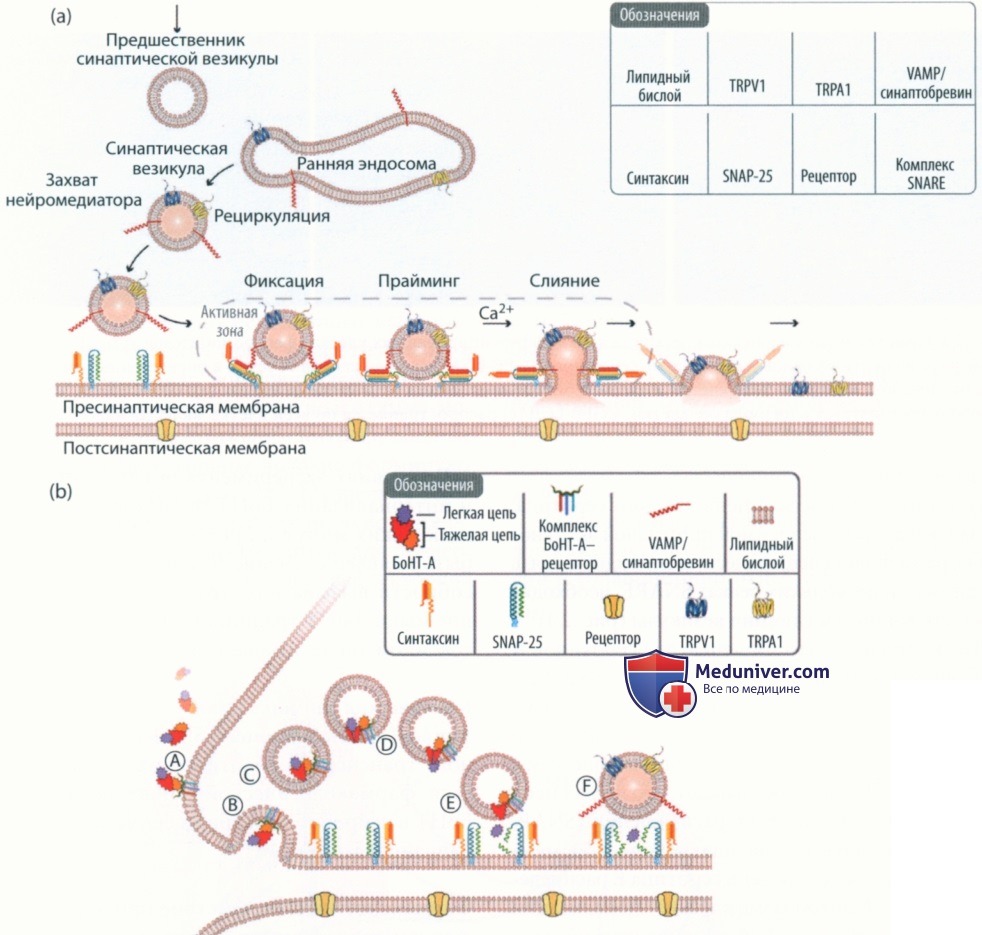
Механизм действия ботулотоксина при мигрени (клиническая фармакология)
Хроническая мигрень характеризуется активацией тригеминоваскулярного пути с центральной и периферической сенсибилизацией с последующим высвобождением таких провоспалительных медиаторов, как субстанция Р, глутамат и CGRP. В активацию периферического пути менингеальными ноцицепторами может быть вовлечено множество рецепторов, включая каналы TRP, рецепторы Р2Х3, чувствительные к АТФ, дофаминовые рецепторы (D1 и D2) и серотониновые рецепторы 5HT1b/1d.
БоНТ-А блокирует высвобождение субстанции Р из культивируемых нейронов спинномозговых ганглиев и стимулируемое, но не базальное высвобождение CGRP из культивируемых нейронов тройничного нерва. Кроме того, в доклинических исследованиях БоНТ-А снижал механическую боль, ингибируя или сокращая экспрессию механочувствительных ионных каналов на поверхности периферических тригеминоваскулярных нейронов. Таким образом, в дополнение к ингибированию/ снижению числа соответствующих рецепторов периферических нервов и ионных каналов при патологии OnaBTX-A может оказывать профилактическое действие при хронической мигрени посредством двойного механизма, включающего блокаду SNARE-опосредованного высвобождения нейромедиаторов воспаления и пептидов из периферических терминалей первичных ноцицептивных афферентных нейронов.
При лечении хронической мигрени OnaBTX-A с профилактической целью вводили в краниоцервикальную область. Благоприятное действие наблюдалось к 4-й неделе, инъекции повторяли каждые 12 недель. Результаты III фазы клинического исследования показали безопасность и эффективность инъекций OnaBTX-A на протяжении 56 нед.. Согласно историям болезней пациентов, получавших до 9 циклов инъекций OnaBTX-A (примерно в течение 2 лет), отмечалось пролонгированное действие препарата в реальной ситуации, выражавшееся в сокращении длительности головной боли.
Рисунок 4. Механизм действия ботулинического нейротоаксина А (БоНТ-A): доставка синаптическими везикулами нейромедиаторов, мембранных каналов и рецепторов: (а) Транспортировка синаптической везикулой (СВ) своего содержимого (например, нейромедиаторов) и билипидной мембраны с ионными каналами и рецепторами. СВ образуют в нервном окончании резервный пул и могут содержать нейромедиаторы. На поверхности большинства СВ присутствуют несколько белков: мембраноассоциированные белковые рецепторы, ванилоидные (TRPV1) и анкириновые рецепторы (TRPA1) из семейства TRP-рецепторов - ионных каналов с транзиторным рецепторным потенциалом. СВ фиксируются на нервном окончании и активной зоне внутренней мембраны, и начинается АТФ-зависимый этап прайминга, в ходе которого в ответ на повышение концентрации Са2+ запускается слияние, экзоцитоз и последующая доставка во внеклеточное пространство не только содержимого СВ, но и липидной мембраны и с белками на поверхность клетки. Для успешного слияния требуется взаимодействие находящегося на мембране везикулы белка VAMP/синаптобревина с расположенными на внутренней мембране белком SNAP-25 с м.м. 25 кДа и синтаксином. В результате образуется комплекс SNARE. В процессе участвуют, но не изображены, и другие белки (например, Muncl8, Rab)41. Мембрана СВ может полностью слиться с терминальной мембраной (полное разрушение и слияние), доставляя таким образом белковые рецепторы (например, TRPV1 или TRPA1) на поверхность клетки. Дополнительный цикл посредством одного из способов эндоцитоза не показан. ОпаВТХ-А расщепляет SNAP-25, препятствуя слиянию СВ и регулируемой доставке рецепторов TRPV1 или TRPA1 к терминальной мембране, снижая тем самым активность рецепторов. Для наглядности СВ с содержимым и липидным бислоем изображены схематично. (b) Механизм действия ОпаВТХ. (А) тяжелая цепь OnaBTX-А связывается с акцепторным комплексом, состоящим из трех компонентов: ганглиозида GT1b, SV2 и рецептора фактора роста фибробластов 3 (FGFR3); (В) интернализация в эндосому и (С) закисление содержимого; (D) конформационное изменение, позволяющее легкой цепи пройти сквозь мембрану эндосомы; (Е) в цитозоле легкая цепь специфически расщепляет SNAP-25 (м.м. 25 кДа), один из реципиентов SNARE-комплекса, необходимых для стыковки СВ с мембраной; (F) расщепление SNARE препятствует слиянию СВ с терминальной мембраной. Это блокирует доставку содержимого СВ (нейромедиаторов) в синаптическую щель, а также доставку карго и экспрессию на поверхности клеток соответствующих рецепторов периферических нервов и ионных каналов.
Имеется как минимум три доказательства того, что БоНТ-А регулирует высвобождение нейромедиатора и количество рецепторов при патологических или стимулируемых, но не при нормальных или базальных состояниях при наличии сенсорного компонента:
1) БоНТ-А блокирует стимулируемый, но не базальный выход CGRP из клеток тройничного нерва, потенциально вызывающий мигрень;
2) БоНТ-А нормализует выделение в уротелиальное пространство АТФ и NO при хроническом повреждении спинного мозга;
3) при нейрогенной гиперактивности детрузора БоНТ-А нормализует концентрацию TRPV1 и Р2ХЗ в мочевом пузыре.
Последнее наблюдение (3) согласуется как с предполагаемой двойной регуляцией конститутивных путей поверхностной экспрессии/проникновения TRP, при которой TRP-каналы достигают плазматической мембраны экзоцитозом от транс-Гольджи или ранних эндосом, так и по регулируемому везикулярному механизму. При последнем рецепторы транспортируются в двойном липидном бислое нейромедиатора или нейропептидов везикул, которые фиксируются и сливаются с мембраной в ходе SNARE-зависимого процесса. В этой модели БоНТ-А блокирует SNARE-регулируемый механизм проникновения, но не конститутивной экспрессии этих сенсорных рецепторов.