Типы наследования в генетике - с точки зрения детского стоматолога
Данная статья на сайте преследует две цели: описать основные принципы генетики и рассмотреть несколько примеров влияния генетических факторов на состояние черепа, лица, полости рта и зубов. Редкие дисплазии, заболевания и синдромы имеют генетическую причину или выраженное генетическое влияние, поэтому все большее внимание уделяется тем генетическим факторам, которые связаны с наиболее распространенными состояниями. Понимание того, как генетические факторы взаимодействуют с экологическими (негенетическими) факторами, влияющими на рост и патологию, приведет к пониманию патогенеза и признанию того, что некоторые группы или индивидуумы могут быть более восприимчивыми или по-разному реагировать на лечение. Дополнительную информацию вы можете найти в разделе генетика сайта - просим пользоваться формой поиска выше.
а) Обзор генетических принципов. Геном представляет собой совокупность генов, содержащихся в наборе хромосом клетки или организма. Гены наименьшие физические и функциональные единицы наследования и находятся в определенных участках хромосом, называемых локусами. Ген может быть определен как ДНК-последовательность, необходимая для синтеза функциональной полипептидной молекулы (продукции белка через промежуточную информационную РНК) или молекулы РНК (транспортной или рибосомной РНК). Генотип обычно определяется набором генов, которые несет человек, и, в частности, относится к конкретной паре аллелей (альтернативным формам определенного гена), которые человек имеет в определенном участке (локусе) своей общей совокупности ДНК, называемой геномом этого человека. Напротив, фенотип — это наблюдаемые свойства и физические характеристики человека, определяемые генотипом индивидуума и окружающей средой, в которой человек развивается в течение определенного периода времени.
Значительные успехи в биохимических методах, используемых для изучения молекулярной биологии клетки и ДНК, поставили исследователей на порог понимания регуляции функций клеток. Не так давно анализ ДНК проводили в мельчайших единицах (пикограммах) ДНК. Это ограничение было необходимо, потому что для исследований в образцах содержалось слишком мало ДНК. Когда исследователи обнаружили фермент ДНК-полимеразу, который мог бы реплицировать ДНК через полимеразную цепную реакцию, данная проблема исчезла. Это продвижение способствовало завершению проекта генома человека, в результате которого было не только определена последовательность нуклеотидов в человеческом геноме, состоящем из перекрывающихся участков, но и постоянно расширяющийся список более чем 1 млн участков изменения последовательности этих нуклеотидов.
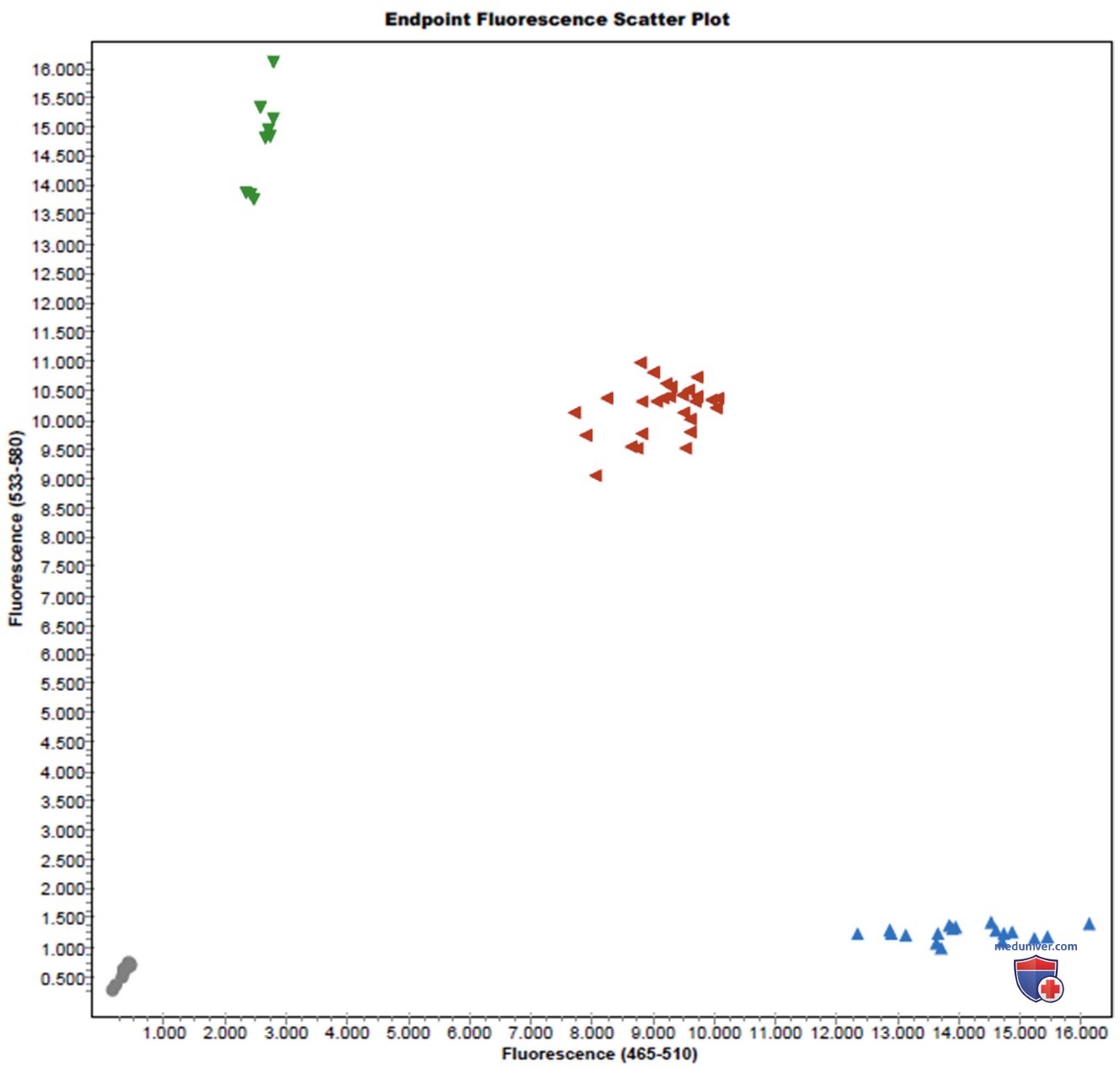
Данные вариации (или полиморфизмы) могут использоваться в качестве маркеров для генетического анализа (включая анализ генетико-экологического взаимодействия) у людей. Геном варьируется от одного человека к другому, чаще всего в пределах единичных изменений ДНК, называемых однонуклеотидным полиморфизмом (SNPs, произносится «снипс»). Основная цель использования карты SNP человека будет заключаться в определении значения тех или иных генов в развитии болезней, имеющих сложную мультифакториальную природу (рис. 1).
Рисунок 1. Генотипирование однонуклеотидного полиморфизма (SNP) на диаграмме рассеяния флуоресцентных точек. Каждый треугольник является индивидуумом, расположенным в области, гомозиготной по одному из вариантов гена (аллели), или гомозиготной по другому варианту (аллели) гена, или гетерозиготной. Генотип индивидуума для данного конкретного SNP можно определить по расположению символа этого человека на участке рассеяния. Например, это график для SNP-гена катехол-о-метилтрансферазы, который важен для восприятия боли. Индивидуумами (обозначены зелеными треугольниками) являются GG, индивидуумы (обозначены красными треугольниками) — AG, синие треугольники — это АА. Серые образцы в левом нижнем углу являются контрольными лицами, которые приняли воду без образца ДНК с целью проверки контаминации компонентов
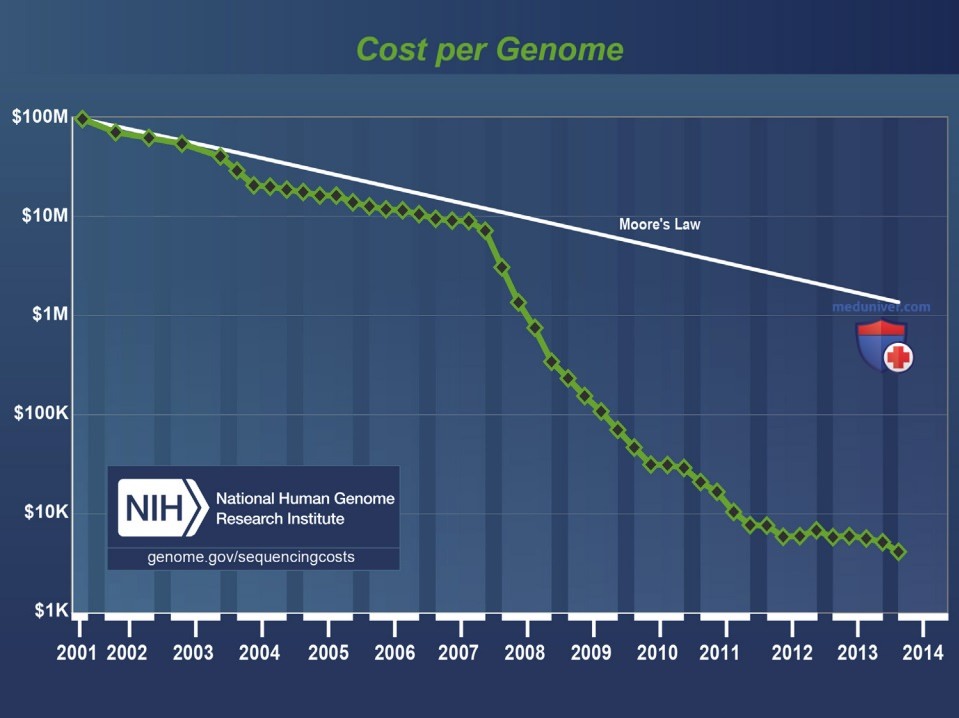
Получение информации о генетической природе некоторых клинических состояний начинает ускоряться из-за того, что называется секвенированием следующего поколения или «глубоким» секвенированием. По сравнению с методами, используемыми в проекте генома человека более 10 лет назад, современные секвенсоры в 50 тыс. раз быстрее, и поэтому пропорционально дешевле (рис. 2). Несмотря на то что затраты снижаются, получение высокого охвата по всему геному (называемое целым геномным секвенированием) по-прежнему дорогостоящее. Таким образом, общепринятой альтернативой служит использование гибридизационного захвата для обогащения образца ДНК только для последовательностей экзонов кодирующих генов (известных как «экзом») приводит к полному экзомному секвенированию.
Рисунок 2. Стоимость секвенирования генома в долларах США оценивается Национальным научно-исследовательским институтом генома человека. По сравнению с гипотетическими данными, отражающими закон Мура, который описывает долгосрочную тенденцию в отрасли компьютерной техники, которая включает в себя удвоение «вычислительной мощности» каждые 2 года, прогресс в снижении затрат на геномное секвенирование был впечатляющим
Определение генов, ответственных за все еще неизвестные менделевские (моногенные) заболевания и признаки, дает немедленную возможность использования этой технологии для полного анализа связи между вариациями генома и фенотипом.
б) Пролиферация клеток и биология развития. Удивительно, но одна оплодотворенная яйцеклетка содержит в себе весь потенциал развития чрезвычайно сложного человеческого организма. Клеточные дифференциалы являются критическим компонентом данного процесса развития, за исключением развития разнообразия антител, что, как правило, происходит в отсутствие генетических изменений или мутаций. Различные типы клеток получают свою специфическую идентичность из определенного подмножества приблизительно 30 тыс. или более генов, присутствующих в геноме.
Типы полипептидов, которые может синтезировать клетка, включают: ферменты, которые катализируют различные процессы гомеостаза клеточного метаболизма; структурные белки, которые образуют внутри- и внеклеточные каркасы, или клеточный матрикс; регуляторные белки, которые передают сигналы от клетки к ядру и модулируют или контролируют специфическую экспрессию гена. В развивающемся эмбрионе клетки находятся в трехмерной среде и реагируют на сигналы от самих себя (аутокринная регуляция), от близлежащих источников (паракринная регуляция) и от анатомически отдаленных источников (эндокринная регуляция). Многие из этих сигналов опосредуются растворимыми молекулами (пептидными или непептидными по происхождению), которые распознаются специфическими рецепторами (белками), расположенными на поверхности или внутри клеток.
В дополнение к сигналам от растворимых факторов клетки могут реагировать на сигналы от клетки к клетке или от клетки к внеклеточной матрице.
Действие «включения» или «выключения» специфических генов, называемое регуляцией экспрессии генов, тщательно спланировано и остается критическим элементом в определении клеточной специфичности и тканевого морфогенеза. Факторы транскрипции связываются с ДНК и либо облегчают, либо подавляют инициирование транскрипции генов, наиболее распространенную контрольную точку экспрессии генов. При развитии черепно-лицевого комплекса все больше доказательств роли семейств генов, содержащих гомеобокс, которые кодируют факторы транскрипции. Впоследствии они имеют решающее значение для контроля сложных взаимодействий между генами, выражающихся во время развития.
Таким образом:
1. Вся генетическая информация заключена в молекуле ДНК. Информация передается от одного поколения клеток к другому посредством репликации или дупликации ДНК.
2. Функция клетки определяется типом РНК, образовавшейся по матрице ДНК. Молекулы РНК управляют синтезом белка в клетке.
3. Факторы транскрипции определяют, какие гены экспрессируются путем продуцирования РНК и последующего белка.
4. Факторы окружающей среды могут влиять на модификацию экспрессии генов путем добавления или удаления химических фрагментов, таких как метильные и ацетильные группы, в ДНК или поддерживающие структуры, называемые эпигенетическими. Если они присутствуют в зародышевой линии, данные изменения могут наследоваться вместе с последовательностью ДНК.
5. Развитие происходит посредством действия специфических факторов транскрипции и других регуляторов продукции белка на специфических генах, которым необходимо экспрессироваться следующими.
в) Хромосомы. ДНК группируется в единицы, называемые хромосомами. У людей 46 хромосом, которые содержат приблизительно 30 тыс. генов, включая многочисленные дубликаты. Из 46 хромосом половыми хромосомами являются X и Y, а остальные 44 хромосомы называются аутосомами. Каждая хромосома имеет пару, называемую гомологом. Все наследственные признаки, за исключением заключенных в X- и Y-хромосомах, контролируются как минимум двумя генами. Таким образом, хромосомный набор человека состоит из 23 пар хромосом (одна пара половых хромосом и 22 пары аутосом).
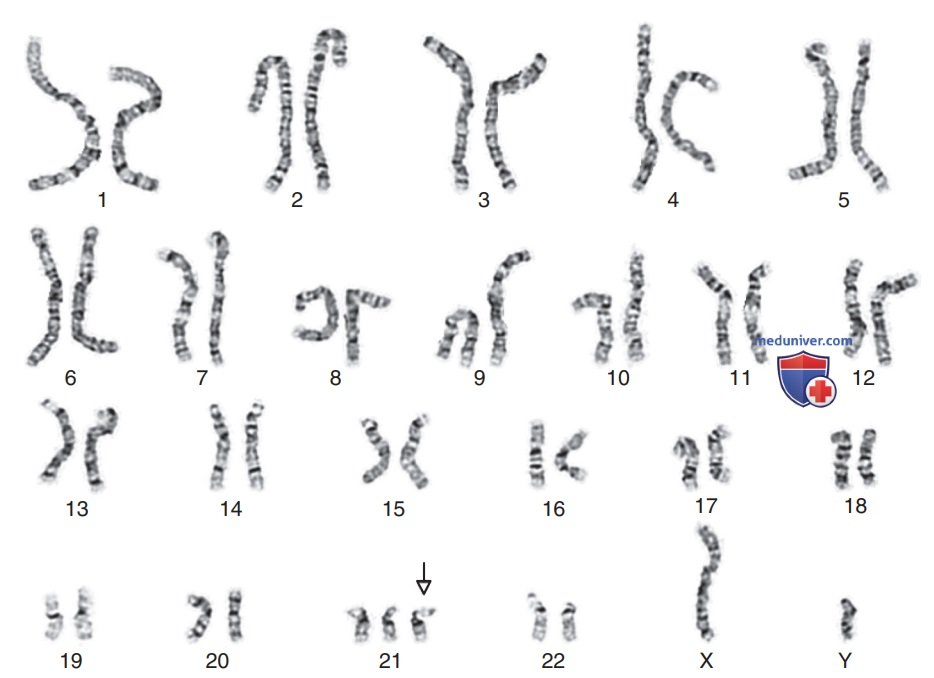
Лежён и коллеги продемонстрировали, что основной причиной синдрома Дауна служит наличие дополнительной специфической хромосомы (21-й) в клетках человека с данным синдромом. Это состояние называется трисомией по данной хромосоме, например трисомия по хромосоме 21 в случае синдрома Дауна. На рис. 3 показан кариотип мужчины с синдромом Дауна. Дополнительная хромосома легко заметна.
Рисунок 3. Кариотип мужчины с трисомией 21-й хромосомы (синдромом Дауна)
Согласно отчету 1959 г., многие заболевания связаны с неправильным комплектом хромосом. Используя сначала микроскопическое исследование структуры хромосом, а затем и ДНК-микрочипирование, было показано, что могут иметь место изменения как самой структуры хромосом, так и их количества. Моносомия аутосом — отсутствие одной аутосомной хромосомы — обычно несовместима с жизнью человека. Моносомия половых хромосом вызывает серьезные нарушения развития наружных и внутренних половых органов, не угрожающие жизни. Наиболее хорошо известным примером моносомии половых хромосом служит синдром Тернера, встречающийся у каждого 5000-го живорожденного. Больные с данным синдромом — женщины, у которых отсутствует одна Х-хромосома. Их кариотип обозначается как 45Х. Другие изменения Х-хромосомы также могут вызвать синдром Тернера. Такие женщины низкого роста, без вторичных половых признаков, бесплодны. В табл. 1 перечислены общие хромосомные аберрации, вызывающие клинические заболевания, включая примеры транслокаций (перенос участка хромосомы на негомологичную хромосому) и делеции (потеря участка хромосомы).
Хромосомные аномалии довольно часто приводят к спонтанным абортам. Около 15% беременностей заканчиваются спонтанными абортами, и причиной половины из них являются хромосомные аномалии. Хромосомные аномалии наблюдаются только у 0,3-0,5% живорожденных и обнаруживаются стандартным микроскопическим кариотипированием. Микроделеции и микродупликации ДНК, не видимые при рутинном хромосомном кариотипном анализе, служат основной причиной развития мальформаций и умственной отсталости. Дополнительный анализ, называемый сравнительной геномной гибридизацией (CGH — comparative genomic hybridization) или хромосомным микроматричным анализом (ХМА), может улучшить диагностическую скорость обнаружения этих небольших хромосомных аномалий. Технология продвинулась настолько, что ХМА теперь является диагностическим тестом первого уровня для людей с множественными врожденными аномалиями, задержкой развития/умственной неполноценностью и расстройствами аутистического спектра, рекомендованным Американским колледжем медицинской генетики и геномики.
Хотя хромосомные микрочипы были разработаны для обеспечения более высокой диагностической значимости, нежели стандартные цитогенетические методы, некоторые страховые компании отказываются предоставлять покрытие для этого теста, утверждая, что он является экспериментальным и не влияет на клиническое ведение пациентов. В отличие от этого утверждения имеются данные, демонстрирующие клиническую значимость ХМА.
д) Наследование признаков в семьях. Наследуемость — это доля общей фенотипической изменчивости в популяции, обусловленная генетической изменчивостью. На индивидуальной основе для бинарного признака (т.е. заболевания или признака, которое индивидуум имеет или не имеет) наследуемость — это не доля заболевания или признака, связанного с генетическими факторами или вызванных ими. Признак с наследуемостью 1, как считают, выражается без какого-либо влияния на окружающую среду, тогда как признак с наследуемостью 0,5 имеет половину изменчивости (от индивидуума к индивидууму) под влиянием факторов окружающей среды и наполовину генотипических факторов. Значения, превышающие 1, также могут возникать, поскольку методология дает оценку наследуемости при некоторых упрощениях, которые могут быть неверными.
Существует общее мнение, что знание наследуемости признака каким-то образом повлияет на то, какое лечение должно проводиться пациенту (например, при малокклюзии — патологии прикуса), или что оно будет определять пределы перемещения зубов или манипуляции с ростом челюсти. Это не так. Способность пациента реагировать на изменения окружающей среды (включая лечение), которая не имеет ничего общего с наследственностью, определяет эти самые пределы. Оценки наследуемости не подразумевают ничего о размере или лимитах размеров, основанных на предполагаемой генетической «предопределенности». Тем не менее оценка наследуемости может свидетельствовать об относительной важности генетических факторов по признаку в группе на тот момент времени. Существует определенная степень генетического влияния на признак — это предварительный шаг к проведению дальнейших специфических исследований генетических связей (с маркерами ДНК) для определения областей генома, которые, по-видимому, связаны с характеристиками данного признака.
Выделяют три типа признаков по количеству контролируемых генов: (1) моногенные, (2) полигонные и (3) мультифакториальные. В последнее время полигонные и мультифакториальные признаки часто объединяются в группу так называемых сложных признаков (реже называемых менделевскими признаками). Моногенные признаки производятся и регулируются одним локусом генов. К моногенным относятся признаки, которые контролируются одним геномным локусом. Такие признаки встречаются среди населения относительно редко (1 из 1000). Однако при поражении контролирующего гена внешний вид человека сразу обращает на себя внимание, например, при альбинизме, ахондроплазии или нейрофиброматозе. Моногенные заболевания встречаются довольно часто у членов одной семьи, их наследование подчиняется законам Менделя.
Полигонные признаки контролируются несколькими геномными локусами. К ним относят такие признаки, как рост, пигментация кожи и умственные способности. Эффекты каждого гена суммируются. Чаще всего это количественные признаки, поэтому они не подчиняются законам Менделя. Важно заметить, что природа этих признаков такова, что они легко подвергаются воздействию окружающей среды в отличие от моногенных признаков. Так, например, очевидно, что окружающая среда может влиять на коэффициент интеллекта (IQ) и рост.
К мультифакториальным относятся признаки, которые тоже контролируются несколькими генами, однако эти признаки возникают только при сочетании влияния нескольких генов с определенными факторами окружающей среды и проявляются при превышении порога предрасположенности. Обычно за такие признаки отвечает большое количество генов, хотя в некоторых случаях они контролируются двумя или тремя геномными локусами. Эффект каждого гена не является добавочным, они действуют только совместно, поэтому признаки нельзя отнести к количественным. Наследование мультифакториальных признаков частично подчиняется законам Менделя. Наиболее распространены такие мультифакториальные наследственные заболевания, как тяжелые пороки развития, например, позвоночная расщелина, дислокация бедренной кости, а также расщелин губы и нёба. Более подробно наследование мультифакториальных признаков будет описано далее.
Изучение наследственных признаков обычно включает выявление специфических признаков у членов семьи и составление родословной. Лицо, с которого начинается составление родословной, называется пробандом. На родословной он указывается стрелкой. Родные братья и сестры пробанда называются сибсами. Клиническое проявление определенного признака, например, цвет глаз или рост, составляет фенотип, а специфический набор генов, ответственный за формирование этого фенотипа, носит название генотип.
Как было сказано ранее, у человека имеется 22 гомологичные пары аутосом и одна пара половых хромосом. Поэтому существует как минимум два гена, локализующихся в одном и том же месте на каждой из гомологичных аутосом, которые контролируют каждый фенотипический признак. Гены, расположенные в одном и том же локусе на паре гомологичных хромосом, называются аллелями. Если оба аллеля в паре идентичны, то говорят о гомозиготном состоянии. Если два аллеля специфического локуса различны, тогда говорят о гетерозиготном состоянии.
Ген, самостоятельно определяющий проявление конкретного признака (в гетерозиготном состоянии), называется доминантным. Если для проявления признака необходимо наличие двух одинаковых генов (гомозиготное состояние), такие гены называются рецессивными. Фактически, доминантным или рецессивным оказывается фенотип, а не сам ген, однако для описания наследования признаков обычно используют термины «доминантный» и «рецессивный» ген.
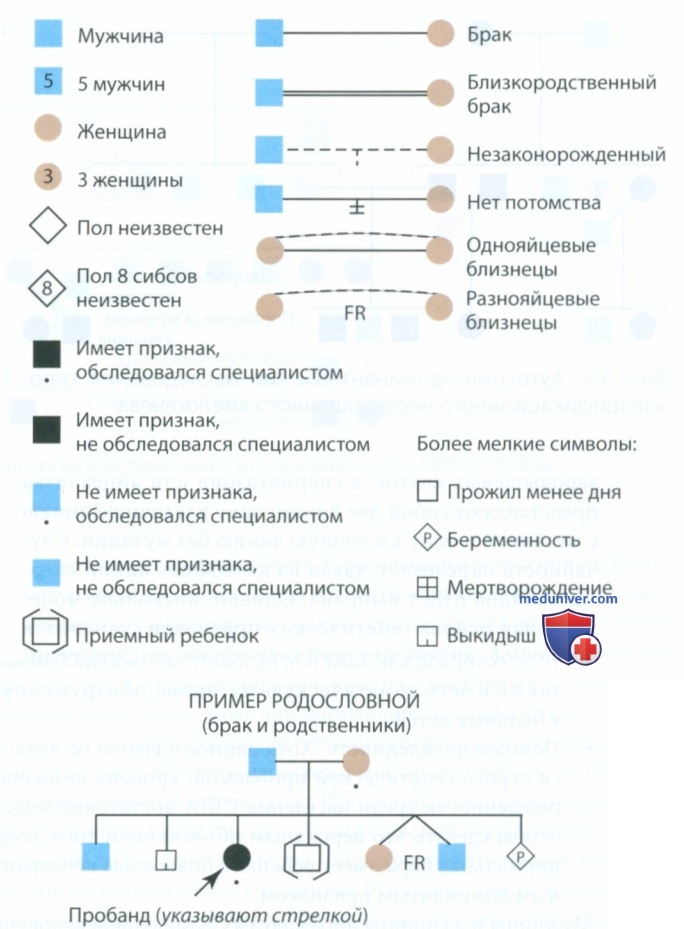
Составление родословной наиболее простой и доступный способ суммирования семейного анамнеза для изучения наследования признаков. На рис. 4 представлены условные обозначения, применяемые при составлении родословной. Тип наследования моногенных признаков определяют три фактора: (1) доминантность или рецессивность, (2) является ген аутосомным или сцепленным с Х-хромосомой и (3) вероятность передачи гена от родителей к потомству с гаметами (сперматозоидом и яйцеклеткой). Важный источник информации о наследуемых признаках — родословная. Каждый стоматолог должен уметь составлять и описывать родословную, так как перед началом лечения наследственного заболевания может потребоваться более точная его диагностика.
Рисунок 4. Условные обозначения при составлении родословных
Далее будут рассмотрены простые типы моногенного наследования. Для иллюстрации основных принципов наследования мы взяли несовершенный амелогенез, так как он может наследоваться по всем менделевским типам.
е) Биология формирования эмали. Тюркер и Шарп представили обзор того, как молекулярные биологи изучают генетические факторы, участвующие в развитии зубов. Клетки нервного гребня, из которых в дальнейшем образуется дентин, и клетки эмалевого эпителия находятся на разных стадиях дифференцировки. Они разделены внеклеточным матриксом. Непосредственное развитие зубов затем зависит от взаимной передачи сигналов от клетки к клетке между этими двумя клеточными слоями. Молекулярные биологи обнаружили структурные белки в эмали и дентине, а также гены, контролирующие развитие этих тканей. Аномалии этих генов обуславливают наличие наследственных нарушений образования эмали и дентина. В настоящее время проводятся исследования, чтобы (1) определить локализацию этих генов и (2) разработать методы биохимической идентификации специфических дефектов, препятствующих нормальному функционированию белков.
В статьях на сайте будут описаны основные генетические принципы наследования на примере наследственных нарушений формирования эмали. Более подробно молекулярная основа наследственных нарушений эмали и дентина описана в отдельной статье на сайте - просим пользоваться формой поиска выше.
На основе клинических, рентгенологических и микроскопических признаков было выделено три типа наследственных дефектов эмали: гипопластический, гипокальцификационный и гипоматурационный.
Эти термины определяют фенотип заболевания. Так, при гипопластическом типе наблюдается снижение количества эмали, которая остается твердой и минерализованной. При этом выделяют два фенотипа: генерализованный (поражается вся эмаль) и локализованный (пятна и борозды определяются на отдельных участках эмали). При гипокальцификационном типе эмалевая матрица повреждается настолько, что ее полноценная минерализация невозможна, в результате чего эмаль становится мягкой и легко подвергается истиранию. При гипоматурацион-ном типе нарушается процесс созревания эмалевых призм. Это происходит после формирования нормальной матрицы, поэтому эмаль имеет нормальную толщину и практически нормальную твердость (небольшая гипокальцификация), однако она пигментирована и ее рентгенологическая плотность снижена.
На основании примера этого заболевания мы можем выделить четыре формы несовершенного амелогенеза, иллюстрирующих четыре основных типа наследования по Менделю: аутосомно-доминантный, аутосомно-рецессивный, Х-сцепленный доминантный и Х-сцепленный рецессивный.
Одна из характерных черт наследственных заболеваний зубов — как временные, так и постоянные зубы поражаются одновременно. Лишь в редких случаях дефект проявляется по-разному в обоих прикусах, например при типе 2 дисплазии дентина. Наиболее часто наблюдается одинаковая клиническая и рентгенологическая картина во временном и в постоянном прикусах. При несовершенном амелогенезе поражаются оба прикуса временный и постоянный.
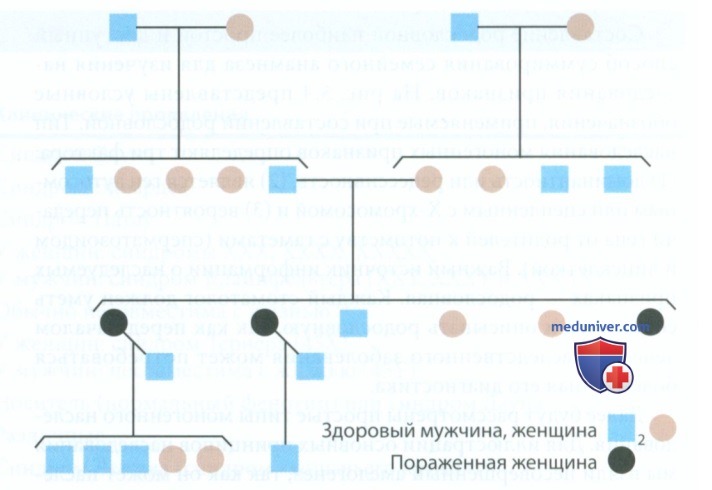
ж) Аутосомно-доминантный тип наследования. Из родословной, которая изображена на рис. 5, можно выделить следующие критерии аутосомно-доминантного типа наследования.
Рисунок 5. Аутосомно-доминантный тип наследования гипокальцификационного несовершенного амелогенеза
1. Наблюдается вертикальный тип наследования, фенотип проявляется последовательно в каждом поколении.
2. В среднем 50% потомства пораженного родителя будут иметь данный признак.
3. У здоровых (без данного признака) родителей рождаются, как правило, здоровые дети.
• Отсутствие пенетратности признака (будет рассмотрено позже).
• Мутация в сперматозоиде или яйцеклетке.
• Герминальный мозаицизм. Данное явление все чаще объясняет подобную клиническую ситуацию. В этом случае один из родителей является мозаичным по линии зародышевых клеток, а сперматазоид или яйцеклетка представляют собой две линии: одну клеточную линию с мутацией и одну клеточную линию без мутации. Случайность определяет, какая из клеточных линий сперматозоидов будет выбрана.
Однако, поскольку молекулярная основа генетических признаков становится очевидной, анализ мутаций может показать, что родитель, который, как кажется, не имеет заболевания, может также иметь соматическую мутацию, обнаруженную у больных детей.
• Незаконорожденность. Хотя данное явление не является строго генетической проблемой, уровень незаконорожденности среди населения США достаточно высок, чтобы сделать это вероятным объяснением того, когда нормальная пара имеет ребенка с полностью пенетрантным доминантым признаком.
4. Мужчины и женщины поражаются с одинаковой вероятностью. Иллюстрацией аутосомно-доминантного типа наследования служит гипокальцификационный тип несовершенного амелогенеза. Для диагностики данного заболевания имеют значение несколько критериев. Во-первых, эмалевая матрица подвержена стиранию; типично скопление большого количества налета на зубах, которые становятся гиперчувствительными из-за обнажения дентина. Во-вторых, на рентгенограмме наблюдаются различная толщина эмали в аппроксимальной области и вид «швейцарского сыра» из-за недостаточной минерализации. Таким образом, тяжелая абразия такой мягкой эмали является обычным явлением.
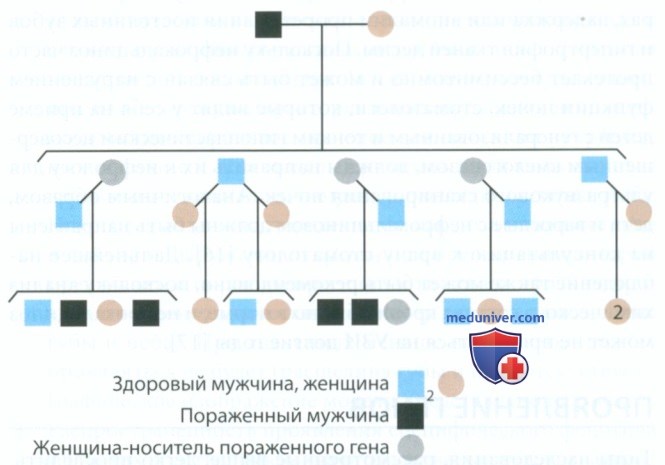
з) Аутосомно-рецессивный тип наследования. При рецессивном типе наследования для проявления признака необходимо наличие двух дефектных генов в одном локусе гомологичных хромосом. Существует три варианта расположения генов: АА — гомозиготный, фенотип не поражен, Аа — гетерозиготный, фенотип не поражен, аа — гомозиготный, фенотип поражен. Чаще всего больной ребенок рождается в том случае, если оба родителя гетерозиготны по данному аллелю (рис. 6).
Рисунок 6. Аутосомно-рецессивный тип наследования пигментного гипоматурационного несовершенного амелогенеза
Рецессивный тип наследования характеризуется следующим.
1. Существует носительство гена. Носитель гетерозиготен по данному рецессивному гену, который фенотипически не проявляется (или имеет незаметные проявления). Родители больного ребенка обычно являются носителями гена (гетерозиготными), они фенотипически здоровы. В некоторых случаях носителей гена можно выявить при помощи специального теста, определяющего наличие одного мутантного гена, например при фенилкетонурии и болезни Тея-Сакса. Это значительно повышает эффективность медико-генетического консультирования.
2. Чем реже встречается рецессивный ген, тем больше вероятность того, что родители больного ребенка — близкие родственники. Если родители больного ребенка гетерозиготны, гомозиготный по рецессивному гену генотип может получиться в одной из четырех возможных комбинаций генов. Таким образом, вероятность рождения больного ребенка в этом случае составляет 25%. Обратите внимание на то, что наблюдается горизонтальный тип передачи признака (поражаются сибсы), а не вертикальный, как при доминантном типе наследования.
Пигментная гипоматурационная форма несовершенного амелогенеза наследуется аутосомно-рецессивно. В этом случае генетический дефект находится в белке, необходимом для созревания эмали на более поздних стадиях формирования зуба. При этом эмаль мягче, чем в норме, однако не такая мягкая и не так подвергается стиранию, как при гипокальцификационном типе. В наружных слоях эмали, которые формируются позднее всего, обнаруживаются коричневая пигментация. Природа этой пигментации неизвестна, однако последняя настолько выражена, что требуется серьезная реставрация зубов. Родословная, демонстрирующая аутосомно-рецессивный тип наследования гипоматурационного дефекта эмали, представлена на рис. 6.
и) Х-сцепленный или сцепленный с полом тип наследования. Гены половых хромосом неодинаково передаются мужчинам и женщинам. Это обусловлено следующими факторами: (1) у мужчин одна Х-хромосома и одна Y-хромосома, а у женщин две Х-хромосомы; (2) гены, находящиеся в Y-хромосоме, ответственны за развитие мужской половой системы. Таким образом, мужчины являются гемизиготными по Х-сцепленным генам, так как у них имеется только половина всех Х-сцепленных генов. А поскольку у женщин имеется две Х-хромосомы, они могут быть гомо- и гетерозиготными по Х-сцепленным генам, также как и по аутосомным генам.
Гемизиготное состояние мужского организма делает возможным образование различных интересных генетических комбинаций. На Х-хромосоме мужчин имеется только один генный локус определенного типа, а для фенотипического проявления рецессивных признаков достаточно наличия лишь одного рецессивного гена. У женщин же для проявления признака Х-сцепленные рецессивные гены должны быть в гомозиготном состоянии (два рецессивных аллеля). Таким образом, Х-сцепленные рецессивные заболевания проявляются практически исключительно у мужчин и крайне редко встречаются у женщин.
Гипопластический несовершенный амелогенез может наследоваться как по аутосомному, так и по Х-сцепленному типу. Далее будет рассмотрен Х-сцепленный тип наследования данного заболевания.
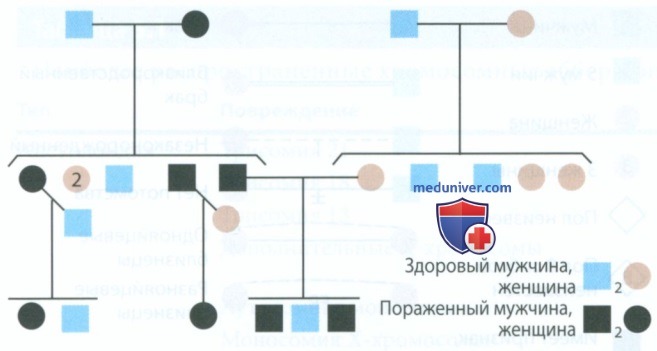
1. Х-сцепленный доминантный тип наследования. На рис. 7 представлена родословная семьи с Х-сцепленным доминантным типом наследования гипопластического несовершенного амелогенеза. Клинические проявления дефекта, особенно у женщин, могут быть очень выраженными.
Рисунок 7. Х-сцепленный доминантный тип наследования гипопластического несовершенного амелогенеза
Опять же, оба зубных ряда могут поражаться одинаково. Дефект поверхности эмали может быть гранулярным, глобулярным или ямочным. Все эти различные фенотипические проявления возникают в результате действия одного гена (или двух аллелей). Эмаль твердая, однако, так как ее слой довольно тонкий, она склонна к скалыванию и патологической стираемости. В некоторых случаях дефект может напоминать гипокальцификационный тип амелогенеза. Однако на рентгенограмме четко видно, что эмаль нормальной плотности, однако толщина ее значительно уменьшена.
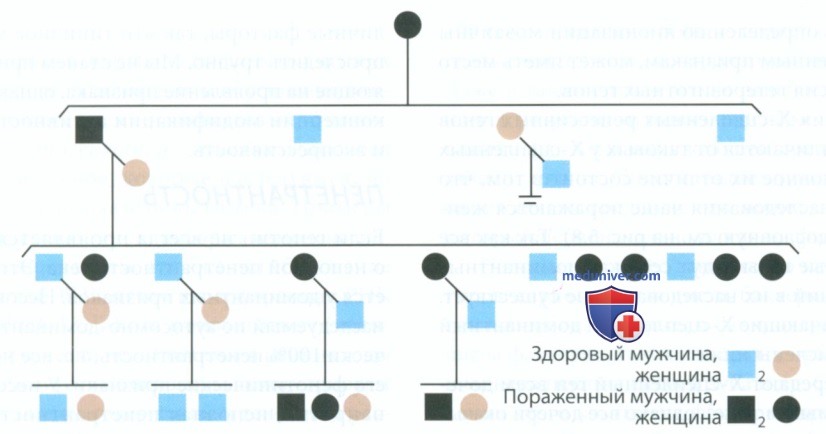
2. Х-сцепленный рецессивный тип наследования. Родословная семьи с Х-сцепленным рецессивным гипома-турационным несовершенным амелогенезом представлена на рис. 8. Генетические критерии диагностики Х-сцепленного рецессивного типа наследования следующие.
Рисунок 8. Х-сцепленный рецессивный тип наследования гипоматурационного несовершенного амелогенеза
1) Так как ген не может передаваться от отца к сыну, у больных отцов не может родиться больной сын. Больной сын может родиться только в том случае, если мать — носитель заболевания.
2) Все дочери больного отца получают Х-сцепленный рецессивный ген. Таким образом, больные отцы через дочерей передают заболевание внукам.
3) Признак проявляется гораздо чаще у мужчин, чем у женщин. Это типично для гемофилии, которая также наследуется по Х-сцепленному рецессивному типу.
Клинические проявления Х-сцепленного рецессивного гипоматурационного амелогенеза обычно имеют следующие признаки: твердость эмали снижена, однако она не мягкая; коронки зубов имеют вид гор, покрытых снегом на вершинах. Отсюда появился термин «снежные» зубы. Рентгенологически эмаль имеет незрелый вид, плотность ее снижена, контрастный переход эмали в дентин отсутствует, даже при нормальной толщине эмали.
Следует отметить, что гетерозиготные самки иногда демонстрируют значительную клиническую экспрессию одного Х-связанного рецессивного гена. Причиной данного очевидного противоречия служит процесс Х-инактивации, называемый лионизацией, после генетика Мэри Лион, которая его обнаружила. Это происходит только у женщин. Все нормальные женские клетки имеют две Х-хромосомы, но большинство генов на одной из двух Х-хромосом инактивируются примерно на стадии развития бластулы. Это приводит к тому, что общее число активных Х-сцепленных генов примерно одинаково у мужчин и женщин. Если самка гетерозиготна по Х-связанному признаку, получаются две популяции клеток.
Одна популяция клеток имеет гены на одной Х-хромосоме, которые активны, тогда как у другой клеточной популяции есть гены на другой Х-хромосоме, которые также являются активными. Когда случайно Х-хромосома с неблагоприятным геном активна в значительной части клеток, ее экспрессия может наблюдаться у этой женщины. Данный случайный дисбаланс происходит не часто, но так как все женщины по определению лионизации мозаичны по отношению к Х-сцепленным признакам, может иметь место фенотипическая экспрессия гетерозиготных генов.
Критерии наследования Х-сцепленных рецессивных генов у мужчин и женщин не отличаются от таковых у Х-сцепленных доминантных генов. Основное их отличие состоит в том, что при доминантном типе наследования чаще поражаются женщины, чем мужчины (родословную см. на рис. 8). Так как все рецессивные Х-сцепленные гены ведут себя как доминантные у мужчин, никаких отличий в их наследовании не существует. Основные критерии, отличающие Х-сцепленный доминантный тип наследования, перечислены ниже.
1. Больные мужчины передают Х-сцепленный ген всем дочерям (как и при рецессивном типе), однако все дочери оказываются больными, так как отец передает свою Х-хромосому дочерям, а Y-хромосому сыновьям.
2. Больные мужчины не могут передать Х-сцепленный ген сыновьям, также как и при Х-сцепленном рецессивном типе.
3. Гетерозиготные по данному признаку женщины передают его 50% своих детей обоего пола, а у гомозиготных женщин будут только больные дети. Последняя ситуация практически не встречается. Поэтому все женщины, имеющие Х-сцепленный доминантный признак, считаются гетерозиготными, если не будет доказано обратное.
Здесь нужно подчеркнуть два момента. Во-первых, наследование Х-сцепленного доминантного гена у женщин не отличается от аутосомного наследования. Поэтому эти два типа наследования можно продифференцировать только при изучении потомства больных мужчин. Во-вторых, Х-сцепленные рецессивные заболевания наблюдаются значительно чаще у мужчин, чем у женщин, а Х-сцепленные доминантные заболевания, наоборот, проявляются в 2 раза чаще у женщин, так как у них вдвое больше Х-хромосом, чем у мужчин.
- Несовершенный амелогенез и синдром нефрокальциноза. Хотя несовершенный амелогенез обычно имеет место в качестве изолированного признака, а не как часть синдрома, многие синдромы включают несовершенный амелогенез в качестве одной из аномалий развития. Редкий, но имеющий большое клиническое значение, эмалево-почечный синдром проявляется несовершенным амелогенезом в совокупности с нефрокальцинозом. Это аутосомно-рецессивное состояние вызвано мутациями в гене FAM20A. В дополнение к гипоплазированной эмали, могут быть обнаружены дентикли в пульпарных камерах, задержка или аномалии прорезывания постоянных зубов и гипертрофия тканей десны.
Поскольку нефрокальциноз часто протекает бессимптомно и может быть связан с нарушением функции почек, стоматологи, которые видят у себя на приеме детей с генерализованным и тонким гипопластическим несовершенным амелогенезом, должны направить их к нефрологу для ультразвукового сканирования почек. Аналогичным образом, дети и взрослые с нефрокальцинозом должны быть направлены на консультацию к врачу-стоматологу. Дальнейшее наблюдение также может быть рекомендовано, поскольку анализ химического состава крови обычно в норме, и нефрокальциноз может не проявляться на УЗИ долгие годы.
к) Проявление генов. Типы наследования, рассмотренные выше, легко проследить, если признак, за который ответственен данный ген, проявляется. К сожалению, на проявление генов оказывают влияние различные факторы, так что типичное моногенное наследование проследить трудно. Мы не станем приводить все факторы, влияющие на проявление признака, однако упомянем две основные концепции модификации активности генов: пенетрантность и экспрессивность.
1. Пенетратность. Если генотип не всегда проявляется в фенотипе, то говорят о неполной пенетрантности гена. Это наиболее часто встречается у доминантных признаков. Несовершенный дентиногенез, наследуемый по аутосомно-доминантному типу, имеет практически 100% пенетрантность, т.е. все носители этого гена имеют его фенотипические признаки. У несовершенного остеогенеза, напротив, неполная пенетрантность, так как исследования показали, что не у всех носителей данного гена проявляется заболевание. Рассмотрим пример наследования расщелины губы и нёба. У деда и внука имеется расщелина губы и нёба, однако у матери мальчика (и дочери деда) эта аномалия отсутствует.
Существует большая вероятность того, что мальчику от деда передалась предрасположенность к дефекту, поэтому у матери не было расщелины. Предрасположенность к данной аномалии можно определить при измерении структур лица. Благодаря значительным успехам в изучении генома человека, мы можем определить локализацию гена, ответственного за данную аномалию, до того, как распознаем его действие на молекулярном уровне и увидим его проявления.
2. Экспрессивность. Если один ген может давать различные фенотипы у пораженных родственников, это говорит о его вариабельной экспрессивности. Пример: несовершенный остеогенез. Признаками данного заболевания служат (1) множественные переломы, (2) голубой цвет склер, (3) несовершенный дентиногенез и (4) отосклероз, приводящий к потере слуха. У членов одной семьи, имеющих данный ген, может наблюдаться один из перечисленных признаков или их сочетание. Минимальной экспрессией гена может быть наличие только голубоватых склер, что часто остается незамеченным. В этом случае можно говорить об отсутствии пенетрантности гена с высокой вариабельностью экспрессивности.
Синдромальные краниосиностозы являются аутосомно-доминантными признаками, связанными с мутациями одного гена. Они также являются хорошими примерами того, как, даже при сильном влиянии одного гена, фенотип может сильно различаться. Несмотря на то что когда-то считалось, что конкретная мутация в данном гене всегда приводит к определенному синдрому, несколько идентичных мутаций в гене рецептора фактора роста фибробластов-2/были обнаружены у пациентов с тремя различными клиническими типами синдромальных краниосиностозов: синдромами Крузона, Пфайффера и Джексона-Вейса.
Другой пример индивидуальной изменчивости подобных аутосомно-доминантных фенотипов с моногенной мутацией произошел, когда у двух индивидуумов в одной и той же семье был классический фенотип синдромов Пфайффера и Аперта. Кроме того, у семи членов семьи были необычно сформированные головы и внешний вид лица, напоминающие синдром Крузона. Фенотип может быть настолько изменчивым, что индивидуум может быть клинически нормальным, но иметь ту же мутацию гена, которая ассоциируется с синдромом Крузона у трех его детей и двух его внуков. Только анализ рентгенологических измерений стал минимальным выражением признаков, свидетельствующих о синдроме Крузона.
3. Эпигенетика. Влияние одного или нескольких модифицирующих генов на их белковые продукты посредством снижения или усиления влияния другого гена в настоящее время называется эпистазом, более старое название — эпигенетика. Эпистаз относится к изменениям в экспрессии генов, которые наследуются, но не вызваны изменением последовательности гена. Примеры эпистаза включают экспрессию генов, которая изменена метилированием или ацетилированием, а также ингибирование экспрессии информационной-РНК путем интерференции РНК или связывания с микроРНК.
Хотя монозиготные (идентичные) близнецы эпигенетически неразличимы в ранние годы жизни, более взрослые монозиготные близнецы демонстрируют значительные различия в общем содержании и геномном распределении 5-метилцитозина ДНК и ацетилирования гистонов, что может создавать различия в экспрессии генов между парами близнецов. Данные эпигенетические факторы могут помочь объяснить взаимосвязь между генетическим фоном человека, окружающей средой, старением и болезнями. Это можно сделать потому, что эпигенетическое состояние изменяется в тканях и в течение жизни, тогда как последовательность ДНК остается по существу одинаковой. Поскольку клетки адаптируются к изменяющейся внутренней и внешней среде, эпигенетические механизмы могут «помнить» эти изменения в нормальном программировании и перепрограммировании активности генов.
Это приводит к новому пониманию того, как взаимодействуют геном и окружающая среда, что может оказать огромное влияние на изучение биологии развития, рака и других заболеваний.
л) Мультифакторальное наследование. Для мультифакториального типа наследования, в отличие от моногенного, характерно, что (1) в проявлении признака принимают участие несколько генов и (2) фенотип возникает в результате совместного действия этих генов. Фенотипические признаки обычно имеют спектр значений (например, рост), а не являются дискретными величинами (признак либо есть, либо отсутствует).
Большинство заболеваний, таких как кариес, не имеют четких критериев наследственной патологии, однако существует определенные параметры, по которым врач может расценивать это состояние как наследуемое.
Мультифакториальное наследование плохо поддается генетическому анализу. Обычно генетики приходят к выводу о наличии мультифакториального заболевания только после того, как исключают моногенный тип наследования. Разработаны специальные методики изучения мультифакториального типа наследования. Наиболее прост из них метод определения сходства между родственниками, согласно которому, чем в более близких родственных связях состоят индивидуумы, тем более схожи у них генетические проявления.
Следует отметить также, что результатом полигенного воздействия являются непрерывные фенотипические вариации. Это имеет большое значение, так как самые распространенные стоматологические заболевания (кариес, заболевания пародонта, аномалии окклюзии) мультифакториальны. Явные только крайние формы вариаций, например множественный кариес у ребенка и отсутствие кариеса у родителей. Нередко эти формы ошибочно принимают за проявления дискретного моногенного признака.
Важно, что мультифакториальные признаки сильно зависят от факторов окружающей среды. Фенотип, контролируемый 100 генами, намного больше подвержен влиянию окружающей среды, чем признак, контролируемый одним или двумя генами. Даже в таком случае это не означает, что на признак, являющийся результатом одного или даже нескольких генов, не могут влиять факторы окружающей среды. Изменение фенотипа зависит от способности индивидуума реагировать на факторы окружающей среды, на что может сильно влиять тот же ген, который первоначально влиял на фенотип.
Например, кариес зависит, по крайней мере, от трех факторов: кариесогенная диета, бактериальная флора и структура тканей зубов, которые, в свою очередь, зависят от большого числа факторов, таких как слюна, зубной налет, формирование и минерализация тканей зуба и др. Очевидно, что в формировании этого комплекса факторов принимает участие большое количество генов. Окружающие факторы, такие как своевременная профилактика фторидами, способны значительно изменить фенотип, не влияя на генотип. Нетрудно представить и другие внешние факторы, которые влияют на развитие кариеса и не имеют отношения к генотипу. Поэтому в большинстве случаев мультифакториальное наследование обуславливает лишь предрасположенность к какому-либо заболеванию, развитие которого зависит от влияния окружающей среды, а также других генетических факторов.
1. Мультифакториальное (сложное) наследование заболеваний человека. При большинстве распространенных заболеваний, таких как сахарный диабет, артериальная гипертензия, а также при многих врожденных аномалиях (расщепление позвоночника, гидроцефалия, расщелины губы и нёба) прослеживается семейная тенденция. Об этом говорит тот факт, что распространенность заболевания среди ближайших родственников выше, чем у остального населения. Однако это соотношение значительно ниже, чем оно могло бы быть при моногенных заболеваниях, и поэтому большинство врожденных аномалий относят к мультифакториальным. Как было сказано ранее, мультифакториальные признаки формируются под влиянием нескольких генов в сочетании с воздействием факторов окружающей среды. Факторы окружающей среды уменьшают или усиливают предрасположенность индивидуума к развитию того или иного состояния.
В отличие от моногенных признаков, мультифакториальные заболевания характеризуются следующими чертами.
1) Каждый человек имеет ту или иную предрасположенность к данному заболеванию, которая определяется совокупным действием генетических факторов и окружающей среды.
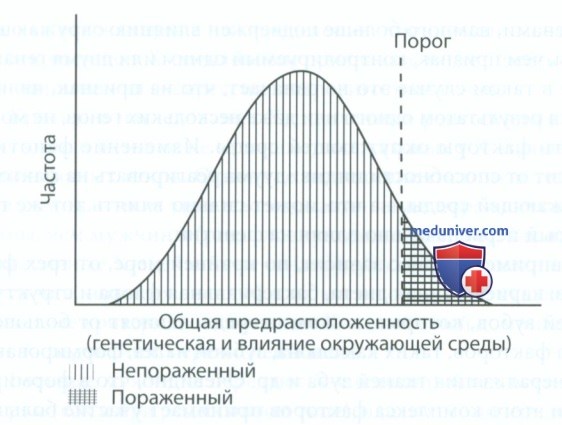
2) Математическим выражением этой предрасположенности служит мультифакториально-пороговая модель, которая представляет собой кривую Гаусса. Как отмечалось ранее, при изучении мультифакториальных признаков необходимо добавить пороговое значение для описания непрерывных величин. Такая модель применима для большинства врожденных аномалий, например расщелины губы и нёба. Пороговая величина определяется как доза влияния генов и факторов окружающей среды, при превышении которой превышается порог экспрессивности гена и признак проявляется фенотипически (например, имеется расщелина губы и нёба). При значениях меньше пороговых признак проявляться не будет (расщелина губы и нёба отсутствует). Графическое изображение модели представлено на рис. 9.
Рисунок 9. Мультифакториальное наследование расщелины губы и нёба
3) Распространенность проявления специфического фенотипа у близких родственников значительно варьирует, что зависит от дозы генов. Например, доза доминантного гена составляет 1 из 2 (50%), а если предположить, что на формирование расщелины губы и нёба влияет несколько факторов, то доза генов снижается как минимум до 1-5%. В действительности данное заболевание встречается даже реже, чем у 1 из 1000. Увеличение дозы гена мультифакториального признака приводит к увеличению распространенности проявления данного признака среди близких родственников. Согласно мультифакториально-пороговой модели большое количество людей, имеющих риск развития заболевания, имеют лишь предрасположенность к нему без клинических проявлений.
В конце XIX в. Гальтон обнаружил, что близнецы могут помочь в изучении взаимосвязи генетических и внешних факторов. В настоящее время интерес к этому методу значительно возрос, так как большинство заболеваний мультифакториальны и подвержены влиянию окружающей среды, а поэтому не поддаются изучению обычными методами. Различают два типа близнецов: однояйцевые (монозиготные или идентичные), которые родились из одной яйцеклетки, оплодотворенной одним сперматозоидом, и разнояйцевые (дизиготные), которые развились из двух яйцеклеток, оплодотворенных двумя сперматозоидами. У монозиготных близнецов одинаковый генотип, а дизиготные близнецы похожи друг на друга не больше, чем братья и сестры, рожденные в разное время. Из этого следует, что различия между однояйцевыми близнецами обусловлены влиянием факторов окружающей среды, а между разнояйцевыми — влиянием как внешних, так и наследственных факторов.
Для применения этого метода необходимо вначале определить, к какому типу относятся близнецы. Если близнецы идентичны по данному признаку (независимо от их зиготности), они считаются согласующимися; если нет — то несогласующимися. Согласованность близнецов обычно выражается в процентном отношении. Так, например, согласованность монозиготных близнецов по расщелине губы и нёба составляет 33%, а дизиготных — 5%.
Другим методом оценки наследуемости признака и оценки доказательств связи фенотипа с полиморфизмами ДНК служит анализ пары сибсов. Выводы о наследуемости могут быть сделаны из вариабельности внутри и между сибсами, количественно определяемой обобщенными линейными моделями. Полиморфные ДНК-маркеры могут быть проверены на генетическую связь (близость) с геном, влияющим на конкретный фенотип, путем проверки того, коррелирует ли величина фенотипической разницы между двумя братьями и сестрами с аллелями, идентичными по происхождению. Аллель считается идентичной по происхождению, если оба сибса унаследовали один и тот же маркерный аллель от одного и того же родителя.
Если маркер связан с геном, вносящим вклад в рассматриваемый фенотип, тогда братья и сестры с аналогичным (если количественным) или одинаковым (если дискретным) фенотипом будут иметь больше идентичных по происхождению аллелей, тогда как братья и сестры с сильно отличающимися фенотипами имеют немного таких аллелей, если какие-либо идентичные по происхождению аллели располагаются вблизи любого гена(ов), влияющего на фенотип.
Кроме того, другой метод поиска маркеров ДНК связан с неустойчивостью связей или анализом ассоциаций. В простейшем понимании это относится к неслучайной ассоциации аллелей в двух или более локусах. Было обнаружено, что некоторые участки ДНК не имеют тенденций к изменению через поколения, в так называемых блоках гаплотипов. Из-за этого тестирование одного однонуклеотидного полиморфизма внутри каждого блока для значительной ассоциации с заболеванием или признаком возможно, когда влияющий локус для этого заболевания или признака находится в блоке гаплотипов или по меньшей мере близко к этому блоку. Частично, поскольку данные блоки гаплотипов встречаются в популяциях, эти анализы могут быть выполнены для не связанных между собой лиц (исследуемых и контрольных), в отличие от анализов сцепления.
Однако если контрольные индивидуумы имеют другой генетический фон, нежели исследуемые, анализ может быть необъективным. Один из способов решения этой проблемы — провести анализ сцепления. Данный анализ вычисляет разницу между значением количественного признака у потомства и средним значением количественного признака у всех потомков во всех исследованных семействах, одновременно рассматривая передачу аллелей от родителя к потомству. Таким образом, в то время как анализ связи пары сибсов включает в себя двух братьев или более и сестер, анализ на сцепление включает в себя родителей и одного или нескольких братьев и сестер.
P.S. Анализ сцепления — это метод генетического картирования, основанный на прослеживании косегрегации генов или генетических маркеров при передаче от родителей к потомкам в ряду поколений.