Фундаментальные принципы геномики - кратко с точки зрения внутренних болезней
Мы вступили в эпоху геномики. Появление новых мощных технологий способствует революции в здравоохранении. Генетическое секвенирование, исходно возникшее как направленное секвенирование одного гена, теперь эволюционировало до параллельного секвенирования нескольких генов.
Такие технологии расширяют возможности выявления генетических причин редких заболеваний, а кроме того, они все чаще и чаще задают направление в лечении и в будущем, вероятно, будут использоваться для диагностики и профилактики таких распространенных заболеваний, как сахарный диабет. В этой и последующих статьях раздела внутренние болезни на сайте обсуждаются фундаментальные основы геномики, базовые принципы новых геномных технологий и способы безопасного применения полученных данных в интересах пациента.
Мы рассмотрим практическое применение геномных технологий в различных областях медицины, включая акушерство, педиатрию, онкологию и инфекционные заболевания, и обсудим, какие изменения будут возможны в здравоохранении благодаря этим технологиям в ближайшие десять лет. Наконец, мы рассмотрим этические аспекты применения этих технологий и их влияние на пациентов и на членов их семей.
а) Укладка генов: дезоксирибонуклеиновая кислота, хроматин и хромосомы. Гены представляют собой функциональные единицы, располагающиеся в дезоксирибонуклеиновой кислоте (ДНК), которая состоит из двух цепей. ДНК образует хромосомы и находится в ядре клетки: ДНК, связанная с мембраной, обнаруживается во всех клетках организма, за исключением эритроцитов и тромбоцитов (рис. 1). ДНК представлена линейной последовательностью, которая образована всего четырьмя азотистыми основаниями: аденином (А), цитозином (С), тимином (T) и гуанином (G).
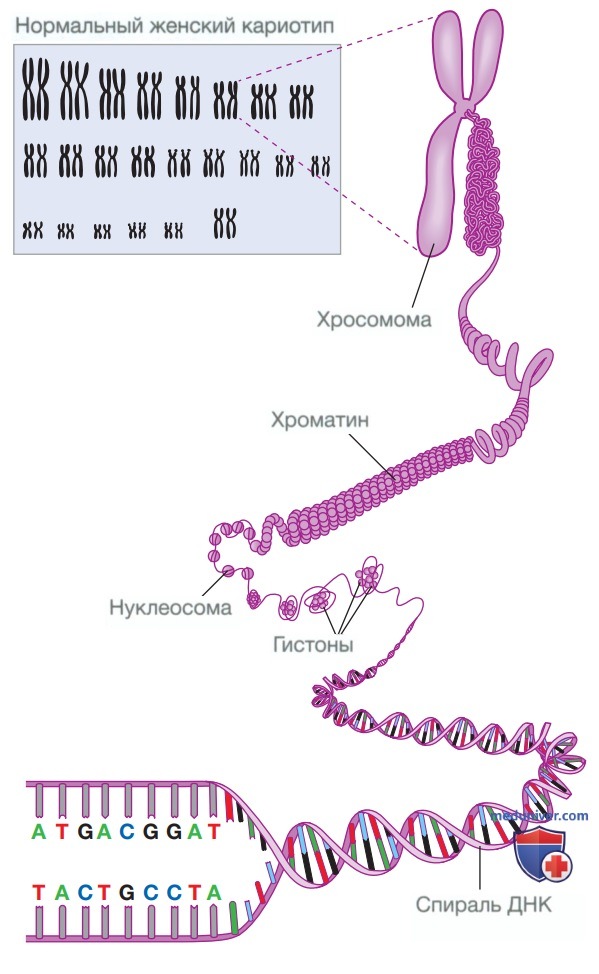
Рисунок 1. Упаковка ДНК, генов и хромосом. Снизу вверх: двойная спираль и комплементарные основания ДНК; хроматин; нормальный женский набор хромосом — кариотип
ДНК образует «двойную спираль» — структуру, напоминающую скрученную лестницу и состоящую из двух комплементарных нитей ДНК, которые соединены водородными связями, локализованными между основаниями противоположных цепей. Причем такие связи могут образовываться только между основаниями С и G и основаниями А и Т. Именно эта особенность ДНК обеспечивает ее точную репликацию и является основой для многих технологий, предназначенных для изучения генома: когда нити двойной спирали ДНК «расходятся», одна из них может выступать в качестве матрицы для создания идентичной нити.
В одной копии генома человека содержится приблизительно 3,1 млрд пар оснований ДНК, накрученных на белки, которые называются гистонами. Единица, состоящая из 147 пар оснований, накрученных на четыре различных белка-гистона, называется нуклеосомой. Последовательности нуклеосом напоминают цепочку бусин. Скрученные и упакованные нуклеосомы образуют хроматин. Плотно уложенный (конденсированный) хроматин называется гетерохроматином, а открытый и менее плотно спирализованный — эухроматином.
В свою очередь, хроматин уложен в хромосомы. Организм человека имеет диплоидный набор хромосом: ядра клеток содержат две копии генома, которые при микроскопии видны в виде 23 пар хромосом, составляющих кариотип. Первые 22 пары хромосом представлены парами одинаковых хромосом и называются аутосомами. Двадцать третья «пара» представляет собой две половые хромосомы: у женщин это две Х-хромосомы, а у мужчин X и Y-хромосомы. В связи с этим нормальный женский кариотип обозначается как 46,XX, а нормальный мужской — как 46,XY.
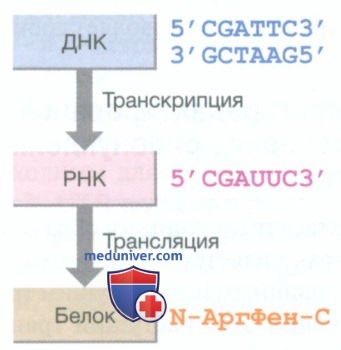
б) От дезоксирибонуклеиновой кислоты к белку. Гены являются функциональными элементами хромосомы, способными передавать информацию с матрицы ДНК путем образования информационной рибонуклеиновой кислоты (иРНК; син.: матричная РНК) с последующим синтезом белка. Геном человека содержит более 20 000 генов, однако многие из них в разных типах клеток остаются неактивными, или «молчащими». Это отражает вариабельность экспрессии генов, отвечающих за характеристики, специфические для конкретного вида клеток. Основой реализации генетической информации является путь, описывающий главные этапы синтеза белка: транскрипцию, сплайсинг, трансляцию и модификацию белка (рис. 2).
Рисунок 2. Основной принцип синтеза белка. Двухцепочечная ДНК является матрицей для синтеза одноцепочечной РНК, которая кодирует аминокислотную последовательность белка. Для каждой из этих цепей характерна направленность: для ДНК и РНК от 5’ к 3’-концу, а для белка — от N-конца к С-концу
В настоящее время такое описание считается чрезмерно упрощенным (в частности, зависимость не всегда является линейной, и один ген может кодировать несколько разных белков), учении синтеза белка.
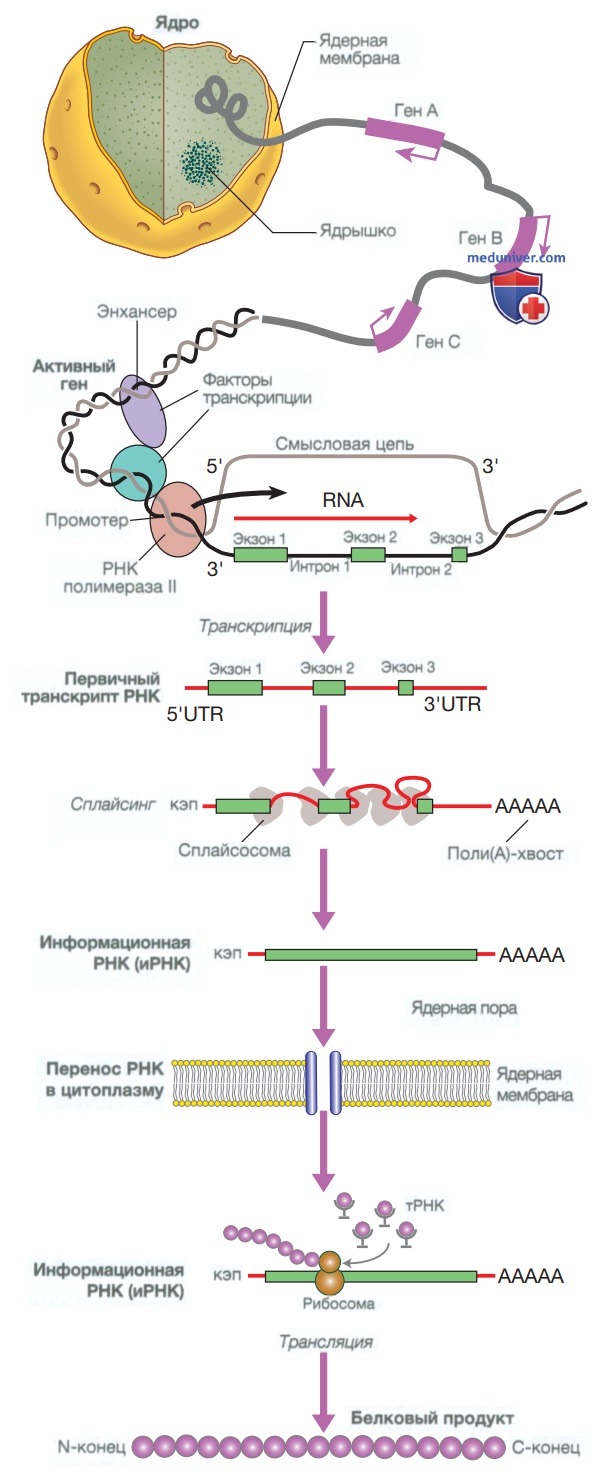
1. Транскрипция: от дезоксирибонуклеиновой кислоты к информационной рибонуклеиновой кислоте. Транскрипция представляет собой синтез рибонуклеиновой кислоты (РНК) на матрице ДНК. Для начала транскрипции фермент РНК-полимераза должен связаться с сегментом ДНК в начале гена, т.е. с геном-промотером. После связывания РНК-полимераза скользит вдоль одной из цепей ДНК и синтезирует молекулу РНК, комплементарную матрице ДНК. Для генов, кодирующих белок, такая РНК является информационной (иРНК). Последовательность ДНК, близкая к концу гена, называется сигналом полиаденилирования и служит сигналом для окончания транскрипции РНК (рис. 3).
Рисунок 3. Синтез РНК и трансляция. Транскрипция гена включает в себя связывание РНК полимеразы II с промотором транскрибируемых генов и другими белками (факторами транскрипции), которые регулируют скорость транскрипции. Первичный транскрипт РНК является копией всего гена и содержит как интроны, так и экзоны. Интроны удаляются в ядре в результате сплайсинга, а экзоны соединяются с образованием информационной РНК. Перед выходом из ядра к 5’-концу РНК («шапка») добавляется метилированный гуанозин, а к З’-концу — цепочка адениновых нуклеотидов («поли(А)-хвост»), Это защищает РНК от расщепления и облегчает ее перенос в цитоплазму. В цитоплазме информационной РНК связывается с рибосомами и образует матрицу для синтеза белка. тРНК — транспортная РНК; UTR (untranslated region) — нетранслируемая область
РНК имеет три основных отличия от ДНК:
• она состоит из одной цепи;
• остатки сахаров в нуклеозидах представлены рибозой, а не дезоксирибозой;
• вместо тимина (Т) РНК содержит урацил (U).
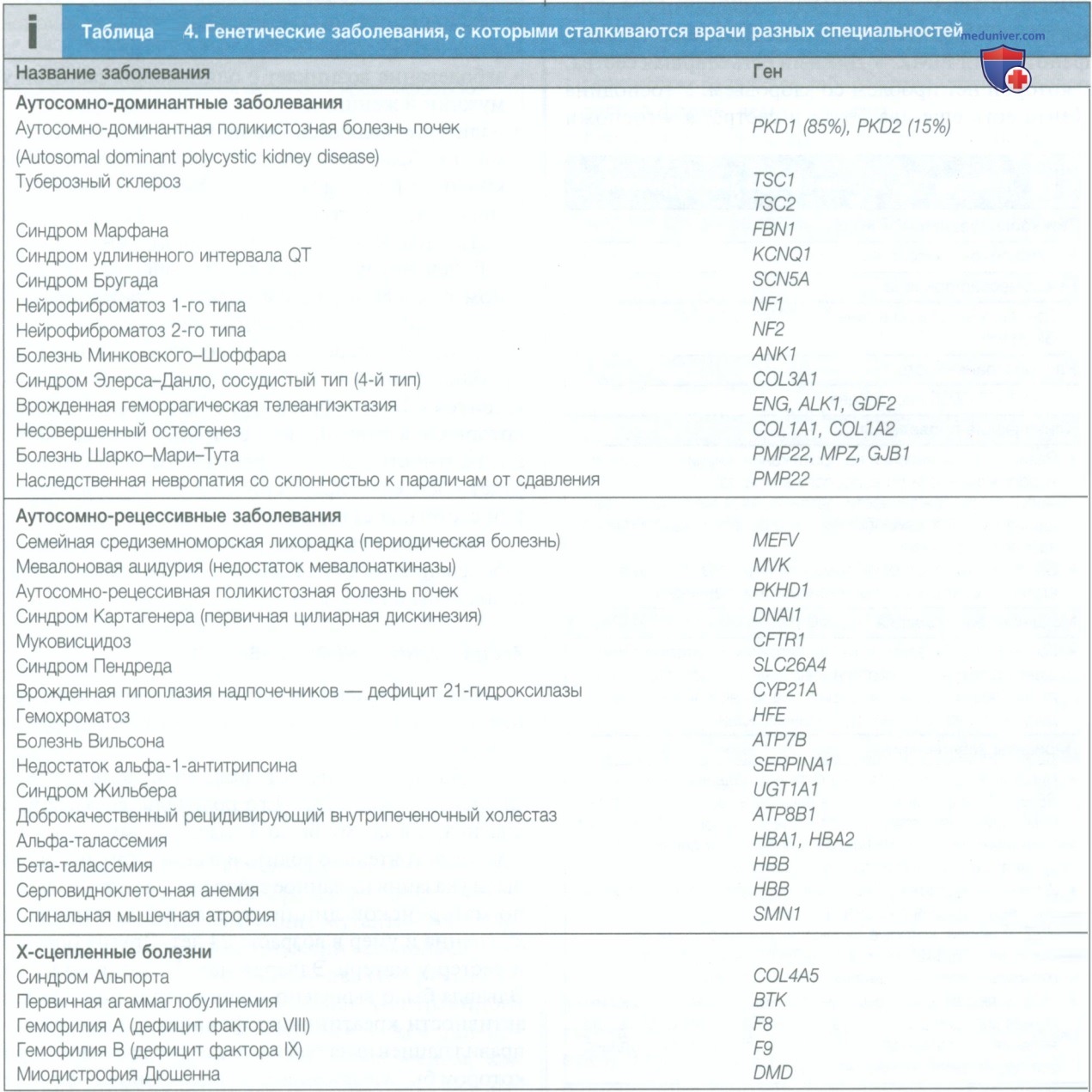
Активность РНК-полимеразы регулируется факторами транскрипции. Эти белки связываются со специфическими последовательностями ДНК в гене-промоторе или с элементами энхансера, которые могут находиться на расстоянии многих тысяч пар оснований от промотора. Петля в хромосомной ДНК приближает энхансер к промотору, в результате чего возможно взаимодействие связанных белков. Геном человека кодирует более 1200 различных факторов транскрипции. Мутации в факторах транскрипции, промоторах и энхансерах могут привести к развитию заболеваний. Например, такая болезнь крови, как альфа-талассемия, обычно вызвана делецией генов (см. табл. 4).
В то же время данная патология может быть результатом мутации в энхансере, расположенном на расстоянии более 100 000 пар оснований (п.о.) от промотора гена α-глобина, что приводит к значительному снижению транскрипции.
На активность или экспрессию генов влияет ряд сложных факторов, взаимодействующих между собой, в том числе доступность промотора гена для факторов транскрипции. ДНК может быть изменена путем добавления метильной группы к молекулам цитозина (метилирование). Если метилирование ДНК происходит на промоторных участках, транскрипция прекращается, поскольку метилцитозины обычно недоступны для связывания с факторами транскрипции. Вторым механизмом, определяющим доступность промотора, является структурная конфигурация хроматина.
В открытом хроматине, который называется «эухроматин», промоторы генов доступны для РНК-полимеразы и факторов транскрипции, поэтому такой хроматин функционально активен с точки зрения транскрипции. В отличие от эухроматина, гетерохроматин плотно упакован и функционально неактивен. Конфигурация хроматина определяется модификацией (в частности, метилированием или ацетилированием) специфических аминокислотных остатков «хвостов» гистоновых белков.
Модификация ДНК и гистонов называется эпигенетической (приставка «эпи-» означает «над» уровнем генома), поскольку она не меняет первичную последовательность кода ДНК, но имеет биологическое значение для функционирования хромосом. Все большее внимание уделяется роли патологических эпигенетических изменений в развитии злокачественных новообразований, поскольку при них становится возможной экспрессия генов, которые в норме являются «молчащими». В результате происходят снижение дифференцированности и пролиферация раковых клеток. Эпигенетические изменения также являются мишенями для терапевтического воздействия.
Например, ингибитор гистондеацетилаз вориностат был успешно использован для лечения кожной формы Т-клеточной лимфомы за счет возобновления экспрессии генов, которые были «молчащими» в опухоли. Эти гены кодируют факторы транскрипции, способствующие дифференцировке Т-клеток, а не их пролиферации, что позволяет добиться регресса опухоли.
2. Сплайсинг, редактирование и расщепление рибонуклеиновой кислоты. В результате транскрипции образуется молекула РНК, которая является копией целого гена и называется первичным или растущим транскриптом. В последующем этот растущий транскрипт подвергается сплайсингу, в результате чего области, не требующиеся для синтеза белка (интроны), удаляются, а сегменты, необходимые в процессе синтеза белка (экзоны), сохраняются и вновь соединяются.
Сплайсинг — это строго регулируемый процесс, который осуществляется многомерным белковым комплексом, называемым сплайсосомой. После сплайсинга молекула иРНК экспортируется из ядра и используется в качестве матрицы для синтеза белка. Многие гены образуют более одной формы иРНК (и, следовательно, белка) в ходе процесса, который называется альтернативным сплайсингом, когда различные комбинации экзонов соединяются вместе. Различные белки, синтезируемые на основании одного и того же гена, могут иметь совершенно разные функции.
Например, в С-клетках щитовидной железы ген кальцитонина образует иРНК, кодирующую ингибитор остеокластов кальцитонин. При этом в нейронах данный ген в результате альтернативного сплайсинга образует иРНК с другим набором экзонов, и данная иРНК кодирует нейротрансмиттер кальцитонин-ген-связанный пептид.
3. Трансляция и синтез белка. После сплайсинга сегмент иРНК, который содержит код, определяющий синтез белкового продукта, называется открытой рамкой считывания. Включение определенной аминокислоты в белок определяется кодоном, который состоит из трех смежных нуклеотидных оснований. В системе есть 64 различных кодона, причем наблюдается некоторая избыточность их количества: 61 кодон кодирует одну из 20 аминокислот, оставшиеся три кодона — кодоны UAA, UAG и UGA (известные как стоп-кодоны) — вызывают прекращение синтеза растущей полипептидной цепи.
Открытая рамка считывания у человека чаще всего начинается с аминокислоты метионина. Во всех молекулах иРНК до и после открытой рамки считывания есть домены, называемые 5’ - и 3’-нетранслируемыми областями (5’-UTR и 3’-UTR) соответственно.
В начале домена имеется кэп-структура, которая защищает иРНК от ферментативного расщепления, другие элементы 5’-UTR необходимы для эффективной трансляции. В домене З’-UTR также содержатся элементы, которые регулируют эффективность трансляции и стабильность иРНК, в частности отрезок адениновых оснований, известный как поли(А)-хвост (см. рис. 3).
В дальнейшем иРНК покидает ядро через ядерные поры и связывается с рибосомами, где происходит синтез белка (см. рис. 3). Каждая рибосома состоит из двух субъединиц (40S и 60S), которые содержат некодирующие молекулы рибосомальной РНК (см. рис. ниже) в комплексе с белками. Во время трансляции с рибосомой связывается другая молекула РНК, известная как транспортная РНК. Транспортные РНК доставляют аминокислоты в рибосому, таким образом сборка синтезируемого белка происходит поэтапно.
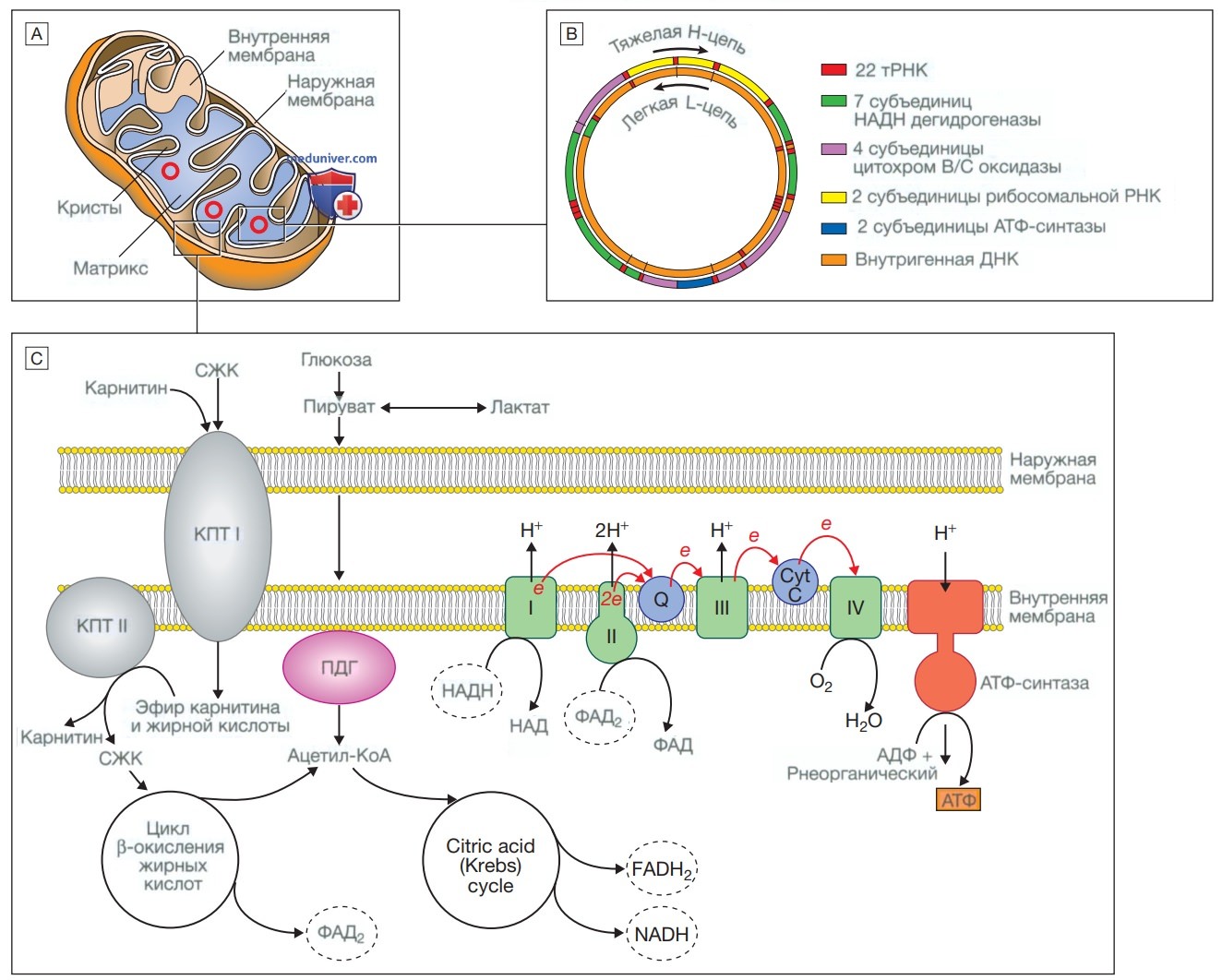
Митохондрия. А — строение митохондрии. Митохондрия имеет две мембраны — гладкую наружную и складчатую внутреннюю, последняя имеет выступы, которые называются кристами. Мембраны формируют два пространства: межмембранное пространство, которое играет важную роль в цепи переноса электронов, и внутреннее пространство (или матрикс), где расположены митохондриальная ДНК и ферменты, участвующие в цикле лимонной кислоты (цикле Кребса) и в цикле β-окисления жирных кислот. В — митохондриальная ДНК. Митохондрия содержит несколько копий кольцевой двухцепочечной ДНК. В ДНК митохондрий есть некодирующий участок и участок, который кодирует гены, отвечающие за образование энергии, молекулы транспортной РНК митохондрий и рибосомальной РНК митохондрий (НАДН — восстановленная форма никотинамидадениндинуклеотида). С — образование энергии в митохондриях. Жирные кислоты переносятся в митохондрию в конъюгированном виде с карнитином, конъюгирование происходит под действием карнитинпальмитилтрансферазы типа 1 (КПТI). Внутри матрикса они расщепляются под действием КПТ II с образованием свободных жирных кислот (СЖК). Последние вступают в цикл β-окисления, в ходе которого образуется ацетил-кофермент А (ацетил-КоА). Пируват может проникать в митохондрию в свободном виде и метаболизируется пируватдегидрогеназой (ПДГ) с образованием ацетил-КоА. Ацетил-КоА включается в цикл Кребса, в результате чего образуются никотинамидадениндинуклеотид и флавинадениндинуклеотид (восстановленная форма). Никотинамидадениндинуклеотид и флавинадениндинуклеотид используются белками в цепи переноса электронов для образования градиента концентрации ионов водорода на внутренней мембране митохондрий. При восстановлении никотинамидадениндинуклеотида и флавинадениндинуклеотида белками I и II соответственно образуются электроны (е), а высвобождающаяся энергия используется для работы протонного насоса с переносом ионов водорода в межмембранное пространство. Кофермент Q10/ убихинон (Q) представляет собой чрезвычайно гидрофобный переносчик электронов, который способен перемещаться в пределах внутренней мембраны. Благодаря переносу электронов белками цепи все большее количество протонов переносится через мембрану до того момента, пока электроны не достигнут комплекса IV (цитохромоксидаза), который использует энергию для восстановления кислорода с образованием молекулы воды. Протонный градиент используется при образовании аденозинтрифосфата под действием фермента аденозинтрифосфат-синтазы, которая состоит из протонного канала и каталитических центров для синтеза аденозиндифосфата из аденозинтрифосфата. При открытии канала ионы водорода попадают в матрикс по градиенту концентрации, при этом происходит выделение энергии, которая используется для образования аденозинтрифосфата
Отдельные молекулы транспортной РНК связывают специфическую аминокислоту и считывают открытую рамку считывания в иРНК посредством «антикодона», который состоит из трех нуклеотидов и комплементарен кодону иРНК (см. рис. 3).
Часть рибосом связана с мембраной эндоплазматического ретикулума, сложной трубчатой структурой, окружающей ядро.
Белки, синтезированные на рибосомах, переносятся в просвет эндоплазматического ретикулума, где происходят формирование укладки и процессинг белка. Из просвета эндоплазматического ретикулума белок переносится в аппарат Гольджи, где он подвергается посттрансляционной модификации, например гликозилированию (ковалентное присоединение остатков сахаров). При этом образуется зрелый белок, который может быть транспортирован в цитоплазму или заключен в пузырьки для дальнейшей секреции.
Клиническую значимость посттрансляционной модификации белков демонстрируют серьезные пороки развития, неврологические заболевания, нарушения гемостаза и изменения мягких тканей, которые наблюдаются при многочисленных врожденных нарушениях гликозилирования.
Посттрансляционная модификация также может быть нарушена в результате синтеза белка с измененными аминокислотными последовательностями. Например, наиболее распространенная мутация при муковисцидозе (AF5O8) приводит к синтезу аномального белка, который не может быть перенесен из эндоплазматического ретикулума и аппарата Гольджи (см. табл. 4).
в) Некодирующие последовательности рибонуклеиновой кислоты. У человека примерно 4500 генов кодируют некодирующие РНК, а не белки. Существуют различные типы некодирующих РНК, в том числе транспортная РНК, рибосомная РНК, рибозимы и микроРНК. МикроРНК, которых насчитывается более 1000, участвуют в посттрансляционной экспрессии генов: они связываются с иРНК, как правило, в домене 3’UTR, что способствует расщеплению определенной иРНК и подавлению транскрипции гена.
В совокупности микроРНК оказывают влияние более чем на половину всех генов человека и играют важную роль в нормальном развитии, появлении злокачественных новообразований и многих дегенеративных заболеваний. В настоящее время микроРНК представляют собой большой исследовательский интерес.
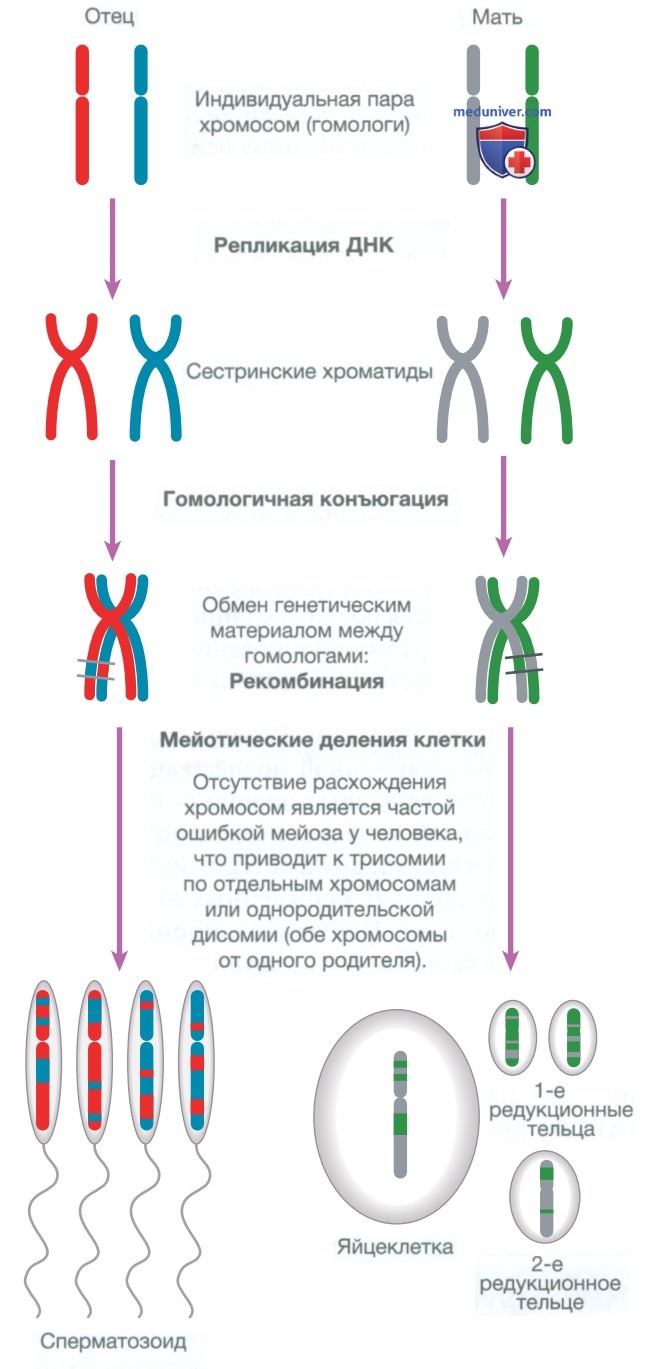
г) Деление, дифференцировка и миграция клеток. В нормальных тканях ряд молекул, таких как гормоны, факторы роста и цитокины, обеспечивает передачу сигнала для активации клеточного цикла — контролируемой программы биохимических событий, которая завершается делением клеток. Деление всех клеток организма, за исключением гамет (сперматозоидов и яйцеклеток, также известных как зародышевая линия), осуществляется митозом, в результате чего образуются две диплоидные дочерние клетки. Сперматозоиды и яйцеклетки делятся путем мейоза, при этом образуются четыре гаплоидные дочерние клетки (рис. 4).
Рисунок 4. Мейоз и гаметогенез: основные хромосомные стадии мейоза у мужчин и женщин. Одна гомологичная пара хромосом изображена разными цветами. Последний этап заключается в образовании гаплоидных половых клеток. В каждом цикле мейоза у мужчин образуется четыре сперматозоида, тогда как у женщин формируется только одна яйцеклетка, а три другие образовавшиеся клетки секвестрируются на периферии зрелой яйцеклетки и образуют периферические редукционные тельца
Стадии деления клеток, не относящихся к зародышевой линии (т.е. соматических клеток), представлены ниже.
• Период клеточного цикла, в течение которого не происходит митоз, называется фазой G0.
• Для перехода к митозу клетка должна пройти подготовительный период, или интерфазу, состоящую из фаз G1, S и G2.
• G1 (интервал 1): синтез клеточных компонентов, необходимых для завершения деления клетки.
• S (синтез): репликация ДНК с образованием идентичных копий каждой хромосомы, которые называются сестринскими хроматидами.
• G2 (интервал 2): исправление ошибок в реплицированной ДНК перед переходом к митозу.
• Митоз (M) включает в себя четыре фазы.
• Профаза: хромосомы конденсируются и становятся видимыми, центриоли движутся к противоположным концам клетки, ядерная мембрана исчезает.
• Метафаза: центриоли завершают миграцию к противоположным концам клетки, а хромосомы, состоящие из двух одинаковых сестринских хроматид, устремляются к экватору клетки.
• Анафаза: нити веретена деления прикрепляются к хромосоме и разводят сестринские хроматиды.
• Телофаза: происходит деконденсация хромосом, вновь формируется ядерная мембрана и образуются две дочерние клетки, каждая из которых имеет по 46 хромосом.
Переход от одной фазы к другой строго регулируется контрольными точками клеточного цикла. Например, контрольная точка между фазой G2 и митозом обеспечивает восстановление всей поврежденной ДНК до разъединения хромосом. Нарушение этих контрольных процессов играет решающую роль в патогенезе злокачественных образований.
• Мейоз — особая форма клеточного деления, характерная для половых клеток (см. рис. 4). Подобно митозу, мейоз состоит из четырех фаз (профазы, метафазы, анафазы и телофазы), но имеет следующие отличия от митоза.
• Мейоз состоит из двух отдельных клеточных делений, известных как мейоз I и мейоз II.
• В результате мейоза число хромосом уменьшается от диплоидного до гаплоидного, что обусловлено наличием тетраплоидной стадии, то есть количество хромосом сначала увеличивается с 46 до 92 (MI S), а при дальнейшем делении уменьшается до 46 (MI М), а затем до 23(МПМ). Таким образом, при оплодотворении яйцеклетки сперматозоидом в образовавшейся зиготе вновь формируется диплоидный набор из 46 хромосом. Уменьшение числа хромосом до гаплоидного происходит в конце мейоза II.
• На тетраплоидной стадии 92 хромосомы представляют собой 23 гомологичные пары сестринских хроматид, между которыми в дальнейшем происходит обмен генетическим материалом. Данный процесс известен как рекомбинация и наблюдается в конце профазы MI. В результате рекомбинации хромосома, которую родитель передает своему потомству, является смесью хромосом, унаследованных родителем от собственных матери и отца.
Отдельные этапы мейоза клеток одинаковы у мужчин и женщин. При этом время, необходимое для деления клеток, очень различается. У женщин мейоз начинается уже во внутриутробном периоде и не завершается до наступления овуляции. Таким образом, одно мейотическое деление клеток может длиться более 40 лет. По мере того как женщина становится старше, расхождение хромосом при мейозе II становится менее эффективным. Именно поэтому из-за отсутствия расхождения хромосом с увеличением возраста матери возрастает риск возникновения трисомии.
У мужчин мейотическое деление не начинается до полового созревания и продолжается в течение всей жизни. Оба мейотических деления в яичках завершаются в течение нескольких дней.
д) Гибель, апоптоз и старение клетки. Все клетки человека, кроме стволовых, имеют ограниченную способность к клеточному делению. Предел Хейфлика — число делений, которое может произойти в культуре в популяции клеток до прекращения деления и перехода в состояние, известное как старение. Эти «биологические часы» представляют большой интерес для изучения нормального процесса старения.
В результате изучения редких заболеваний, связанных с преждевременным старением (прогерические синдромы), стала известна важная роль механизмов репарации ДНК в процессе старения. Например, при синдроме Вернера мутация ДНК-геликазы (фермента, который разделяет две цепи ДНК) приводит к невозможности репарации ДНК и преждевременному старению.
Четкий механизм гибели клеток наблюдается при апоптозе, или запрограммированной гибели клеток.
Апоптоз является активным процессом, который происходит в нормальных тканях и играет важную роль в формировании, ремоделировании тканей и иммунном ответе. Сигнал, который запускает апоптоз, специфичен для каждой ткани или типа клеток. Этот сигнал активирует ферменты каспазы, которые активно разрушают компоненты клетки, включая хромосомную ДНК. Такой распад приводит к гибели клеток, однако погибшая клетка содержит характерные пузырьки, называемые апоп-тозными тельцами. Фагоцитарные клетки иммунной системы, в частности макрофаги, распознают и удаляют погибшую клетку, не вызывая при этом воспалительный ответ.
Третий механизм гибели клеток — некроз. Это патологический процесс, при котором клеточное окружение теряет один или несколько компонентов, необходимых для жизнедеятельности клеток. Самой распространенной причиной некроза, по-видимому, является гипоксия.