Классы генетических вариантов - кратко с точки зрения внутренних болезней
Существуют различные классы изменений человеческого генома, которые формируются в зависимости от размера вовлеченного сегмента ДНК и/или механизмов, приводящих к изменениям.
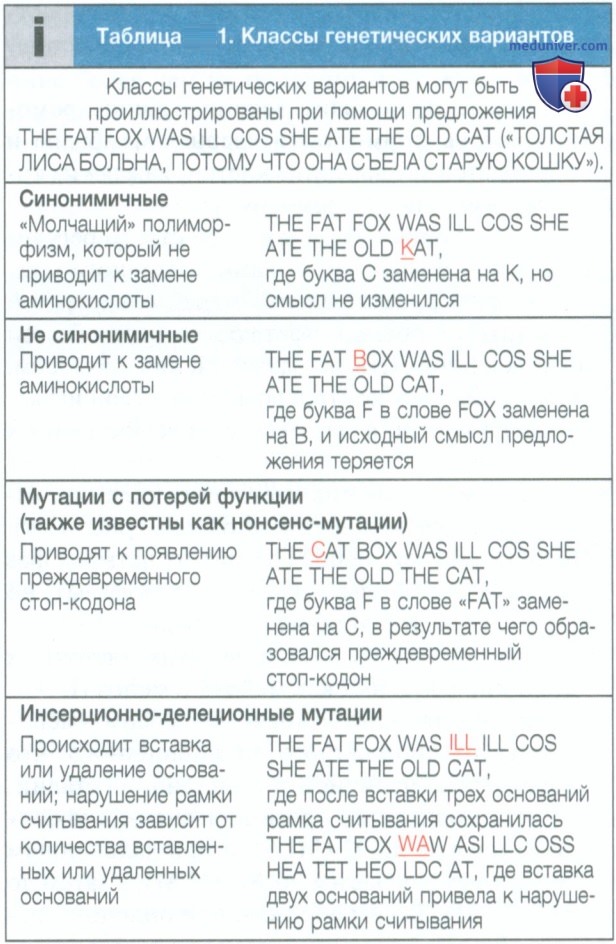
а) Нуклеотидные замены. Замена одного нуклеотида другим является наиболее частым типом вариации генома. В зависимости от частоты и функциональных последствий эти изменения известны как точечные мутации или однонуклеотидные полиморфизмы. Они возникают вследствие ошибочного включения нуклеотида во время синтеза ДНК или вследствие химической модификации основания. В случаях, когда эти замены возникают в пределах открытой рамки считывания белка, который кодируется геном, они дополнительно классифицируются как:
• синонимичные — вызывают изменения кодона, которые не приводят к изменению аминокислоты;
• не синонимичные (также известны как миссенс-мутации) — приводят к изменению кодона и кодируемой аминокислоты;
• мутации с потерей функции (или нонсенс-мутации) — появление преждевременного стоп-кодона и как следствие укорочение белка;
• сплайсинговые мутации — происходят в сайтах сплайсинга, которые чаще всего располагаются на границе интрона и экзона.
Указанные типы мутации представлены в табл. 1, а их примеры приведены на рис. 1 и 2.
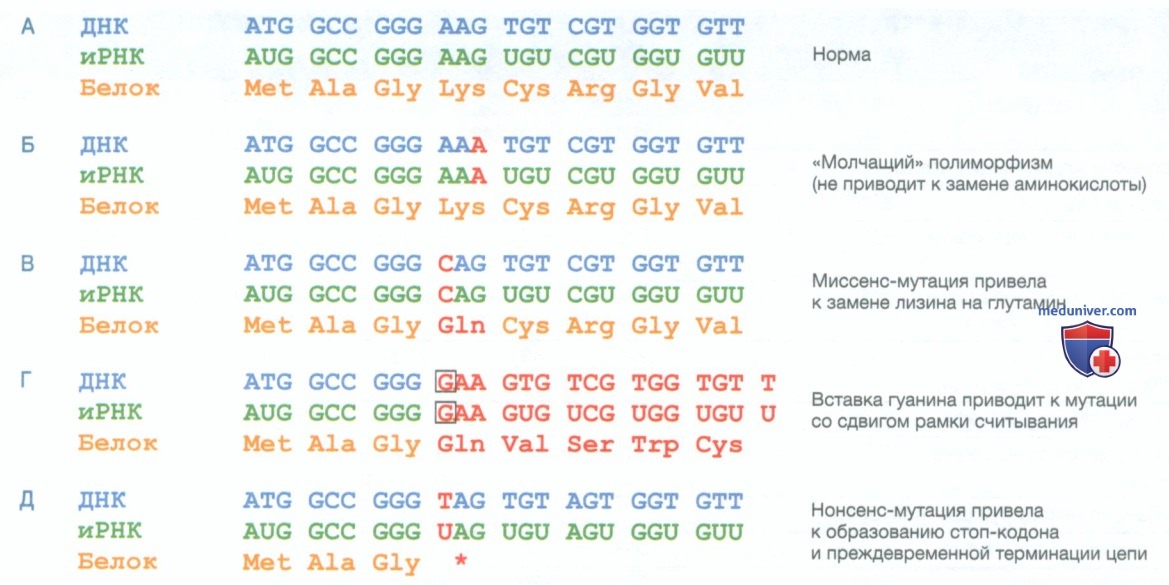
Рисунок 1. Различные типы мутаций, затрагивающих кодирующие экзоны. А — нормальная последовательность. Б — синонимичная нуклеотидная замена в третьем основании кодона, образующаяся аминокислотная последовательность остается неизменной. В — миссенс-мутация, при которой нуклеотидная замена приводит к замене одной аминокислоты (AAG, кодирующий лизин, на CAG, кодирующий глютамин). Г — инсерция остатка G (обозначена рамкой) вызывает мутацию со сдвигом рамки считывания, полностью изменяя аминокислотную последовательность в 5’-3’ направлении. Это обычно приводит к мутации с потерей функции. Д — нонсенс-мутация, приводящая к замене одного нуклеотида в кодоне лизина (AAG) с образованием преждевременного стоп-кодона (TAG)
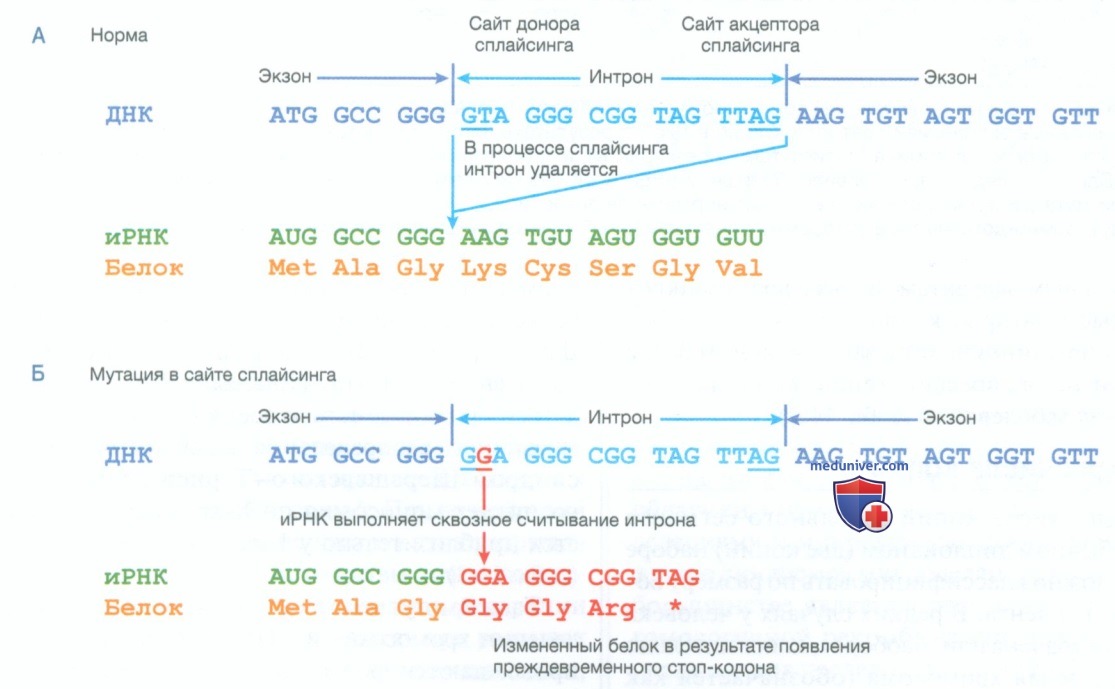
Рисунок 2. Мутации сайта сплайсинга. А — представлена нормальная последовательность, содержащая два экзона и промежуточный интрон (обозначен синим цветом) с сайтом донора (AG) и сайтом акцептора (GT) сплайсинга. Обычно интрон удаляется в результате сплайсинга с образованием зрелой информационной РНК, которая кодирует белок. Б — при мутации сайта сплайсинга происходит мутация сайта донора. В результате этого сплайсинг больше не происходит, что приводит к сквозному считыванию интрона, который содержит преждевременный стоп-кодон, в 5’-3’ направлении после мутации
б) Инсерция и делеция. Один или несколько нуклеотидов могут быть введены в последовательность ДНК или удалены из нее, что приводит к полиморфизму или мутации в виде инсерции/делеции (инсерционно-делеционная мутация или полиморфизм) (см. табл. 1 и рис. 1). Если задействованное количество нуклеотидов кратно трем, происходит мутация внутри рамки считывания. Если инсерция или делеция затрагивает один или два нуклеотида в пределах открытой рамки считывания гена, кодирующего белок, могут возникнуть серьезные изменения, поскольку при этом нарушается тройная нуклеотидная последовательность кодонов и появляется мутация со сдвигом рамки считывания. Воздействие таких мутаций, как правило, серьезное, поскольку аминокислотная последовательность полностью нарушена.
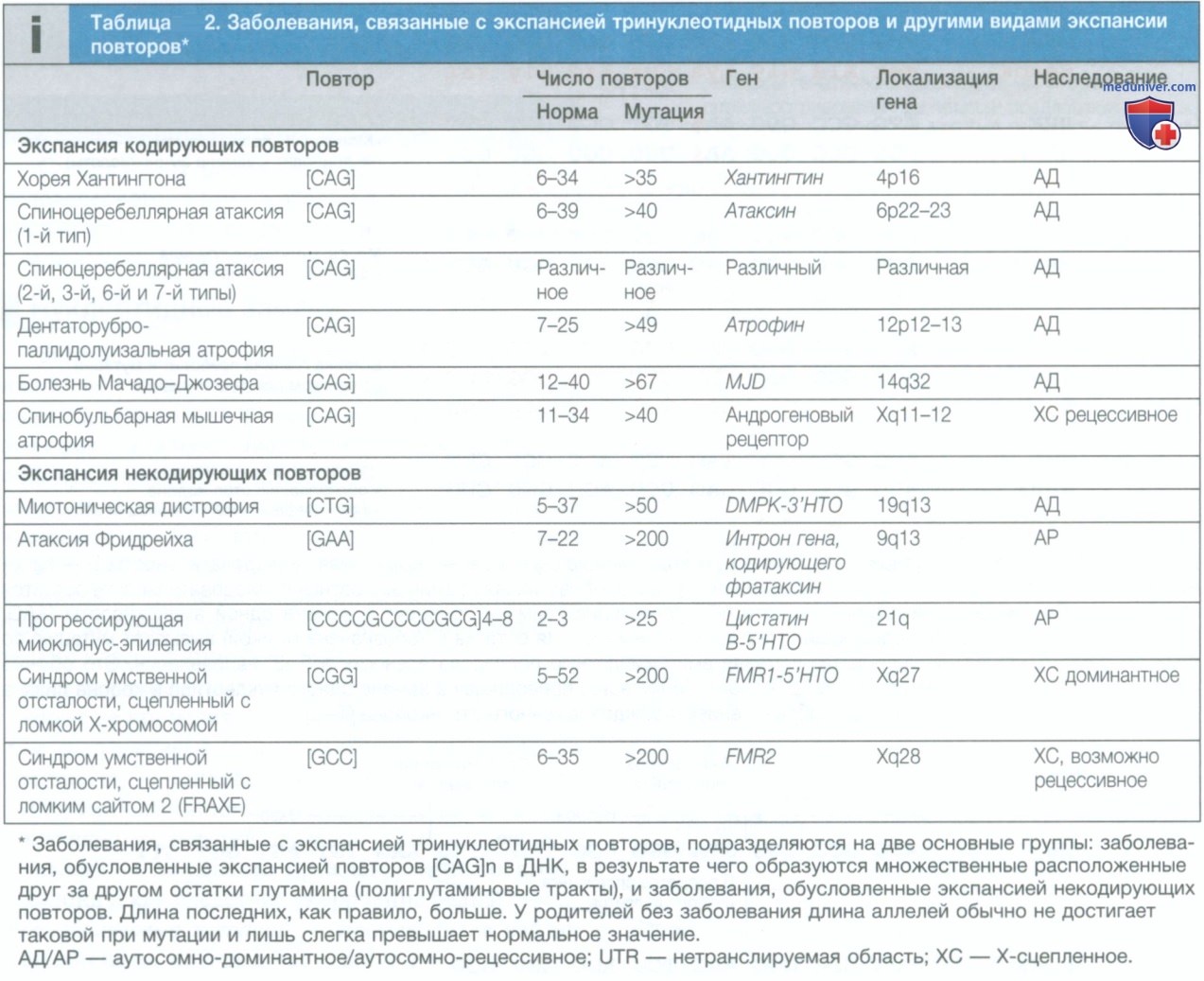
в) Мутации в простых тандемных повторах. Считается, что изменения длины простых тандемных повторов ДНК возникают в результате «проскальзывания» ДНК во время мейоза и называются микросателлитными (малыми) или минисателлит-ными (более крупными) повторами. Такие повторы нестабильны и могут увеличиваться или уменьшаться в разных поколениях. Эта нестабильность пропорциональна размеру исходного повтора, т.е. более длинные повторы, как правило, более нестабильны. Многие микросателлитные и минисателлитные повторы встречаются в интронах или в хромосомных областях между генами и не обязательно приводят к нежелательным эффектам.
В то же время микро-сателлитные повторы, которые влекут за собой дублирование аминокислот в затрагиваемом белке или влияют на экспрессию генов, вызывают ряд генетических заболеваний (табл. 2).
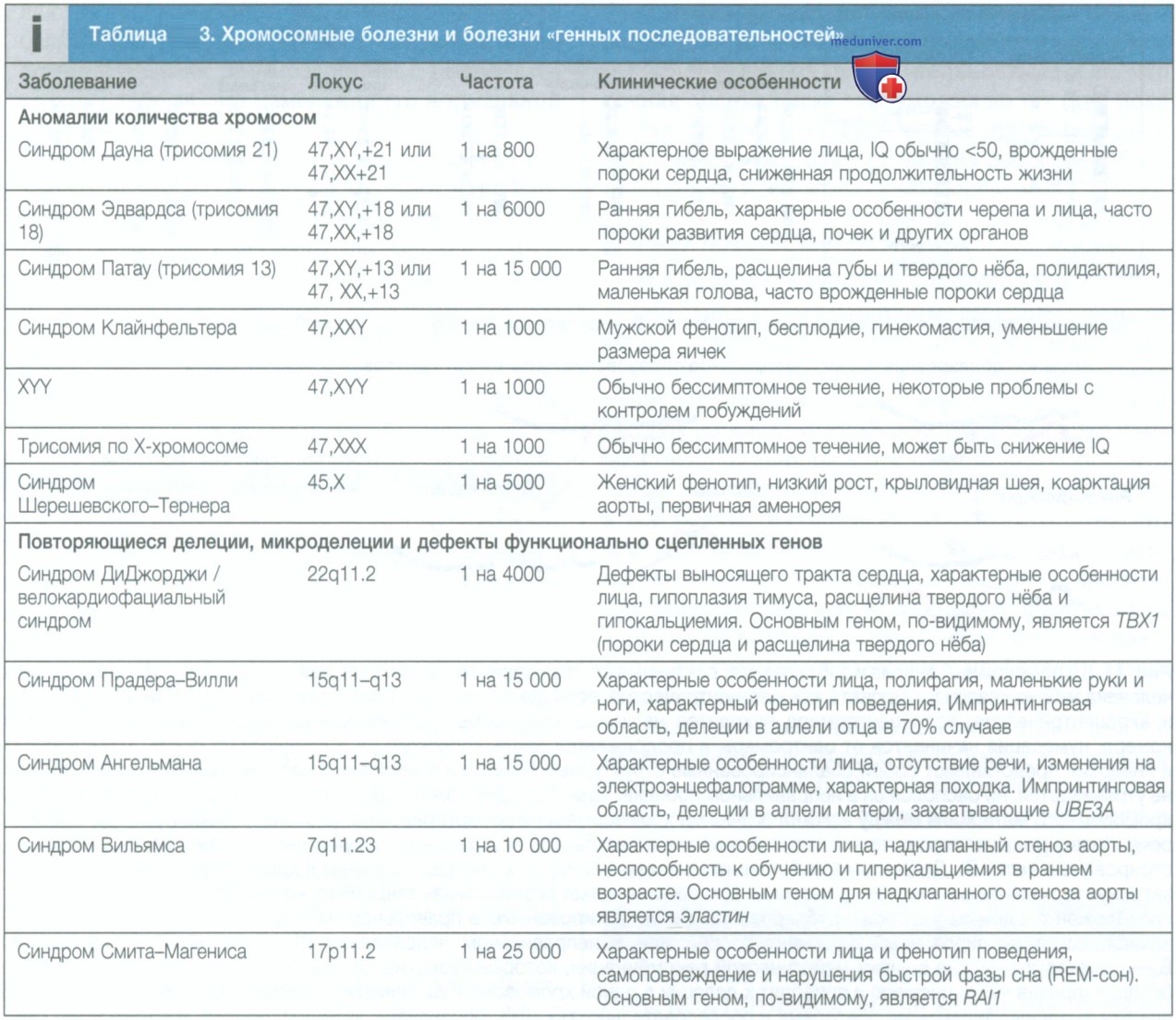
г) Вариации числа копий. Вариации числа копий отдельного сегмента генома в обычном диплоидном (две копии) наборе хромосом можно классифицировать по размеру вовлеченного сегмента. В редких случаях у человека может быть утрачена или, наоборот, дополнительно появляется целая хромосома (обозначается как моносомия и трисомия соответственно). Такие аномалии числа хромосом чаще всего возникают в результате процесса, который называется отсутствием расхождения, когда пары гомологичных хромосом во время мейоза II не разделяются. Наиболее частыми видами трисомии являются синдром Дауна (трисомия 21), синдром Эдвардса (трисомия 18) и синдром Патау (трисомия 13).
Моносомии аутосом (которые есть во всех клетках, в отличие от мозаичного распределения) не существует, однако синдром Шерешевского—Тернера, при котором возникает моносомия по Х-хромосоме, выявляется приблизительно у 1 из 2500 живорожденных (табл. 3).
Также могут возникать большие инсерции или делеции хромосомной ДНК, которые обычно сопровождаются формированием неспособности к обучению и/или врожденных пороков развития. Такие структурные хромосомные аномалии обычно формируются в результате одного из двух различных процессов:
• негомологичное соединение концов;
• неаллельная гомологичная рекомбинация.
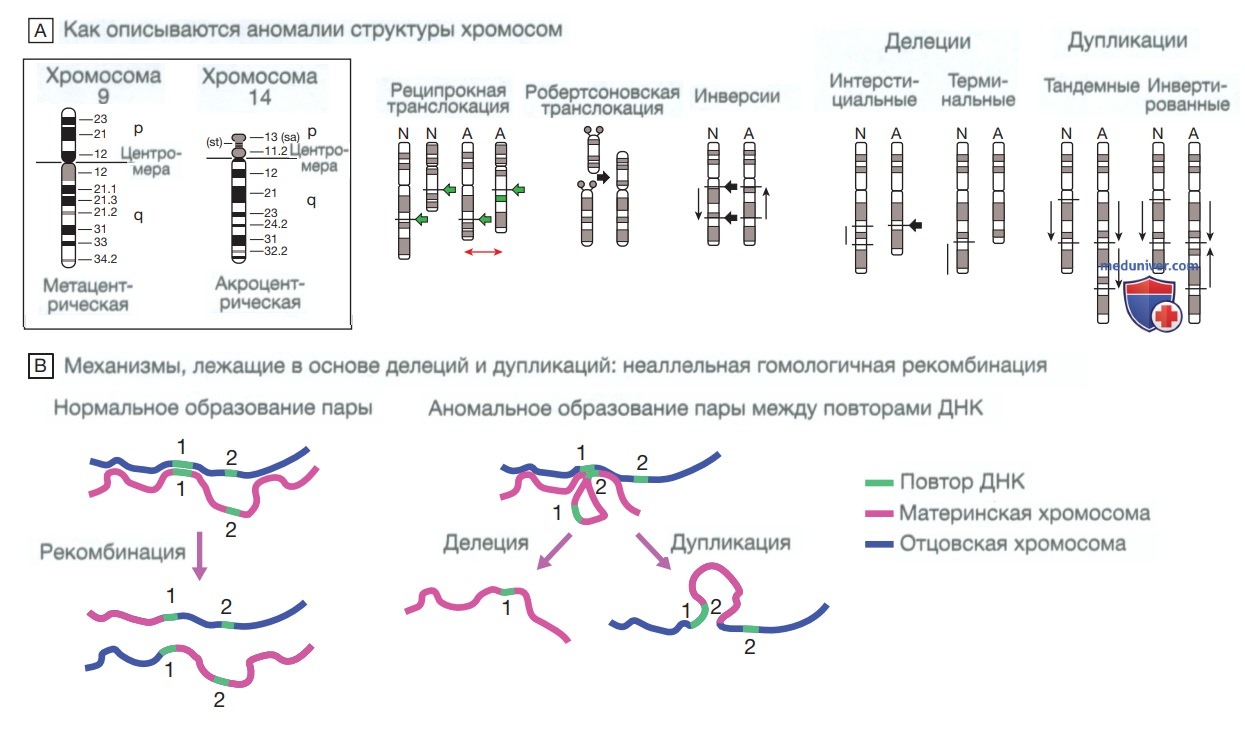
Случайные двухцепочечные разрывы в ДНК представляют собой необходимую часть мейотиче-ской рекомбинации и также происходят во время митоза с предсказуемой частотой. При воздействии ионизирующего излучения частота этих разрывов резко возрастает. Обычно разрывы точно восстанавливаются с помощью механизмов репарации ДНК внутри клетки. Однако иногда при разрывах сегменты ДНК, которые в норме не соприкасаются, оказываются соединены («негомологичное соединение концов»). При соединении фрагментов разных хромосом происходит транслокация. Если фрагменты принадлежат одной и той же хромосоме, возникают инверсия, дупликация или делеция фрагмента (рис. 3). Крупные инсерции, вставки и делеции могут быть видны цитогенетически в виде хромосомных делеций или дупликаций.
Рисунок 3. Хромосомный анализ и заболевания, связанные со структурными хромосомными нарушениями. А — хромосомы человека можно классифицировать как метацентрические, если центромера расположена ближе к середине хромосомы, и акроцентрические, если центромера находится на конце хромосомы. Дискам каждой хромосомы присваиваются номера, нумерация начинается от центромеры и продолжается вдоль короткого (р) и длинного (q) плеча. Транслокации и инверсии представляют собой сбалансированные структурные аномалии хромосом, в которых генетический материал не утрачивается, но расположен в неправильном порядке. Транслокации можно разделить на реципрокные (прямой обмен хромосомного материала между негомологичными хромосомами) и робертсоновские (слияние акроцентрических хромосом). Делеции и дупликации также могут возникать в результате неаллельной гомологичной рекомбинации (продемонстрировано в части В). Делеции могут быть интерстициальными, если они расположены в хромосоме, и концевыми, если затронут терминальный участок хромосомы. Дупликация может возникать как тандемная, когда дублированный фрагмент расположен следом за участком, дублированным и ориентированным в правильном направлении, либо как инвертированная, когда дублированный фрагмент расположен в неправильном направлении (Н — норма; П — патология). В — существуют частые ошибки мейотической рекомбинации, которые известны как неаллельная гомологичная рекомбинация (правая часть рисунка) и приводят к делеции в одной хромосоме и дупликации в гомологичной хромосоме. Такие ошибки вызваны тандемными повторами в последовательностях ДНК (обозначены зеленым цветом), которые могут смещаться и связываться друг с другом, тем самым «обманывая» ДНК и заставляя воспринимать, что образование пар до рекомбинации является правильным
Если аномалии слишком малы, чтобы их можно было обнаружить при микроскопии, их называют микроделециями и микродупликациями. Описано множество синдромов микроделеции, которые в своем большинстве являются результатом неаллельной гомологичной рекомбинации между повторами очень схожих последовательностей ДНК. Это приводит к рецидивирующим хромосомным аномалиям и клиническим синдромам, которые возникают у людей, не являющихся родственниками (рис. 3 и табл. 3).
д) Последствия геномных вариаций. Последствия отдельной мутации зависят от многих факторов, включая тип мутации, природу белка и положение варианта в белке. Мутации могут оказывать существенное или незначительное воздействие на функции генов и клеток. «Классические» генетические заболевания обусловлены наличием вариаций, приводящих к значимым последствиям, тогда как вариации со слабовыраженным действием могут играть роль в патогенезе распространенных заболеваний, имеющих генетический компонент (например, сахарный диабет).
• Нейтральные варианты не влияют на качество или тип образующегося белка.
• Мутации с потерей функции приводят к потере или снижению нормальной функции белка. Архетипичными мутациями с потерей функции являются делеции всего гена, однако мутации с потерей функции или инсерционно-делеционные мутации (в начале открытой рамки считывания), миссенс-мутации, влияющие на критический домен, и мутации сайта сплайсинга также могут приводить к потере функции белка.
• Мутации с приобретением функции приводят к усилению функции белка. Они, как правило, являются несинонимичными, изменяют структуру белка и активируют/изменяют его нормальную функцию за счет взаимодействия с новым субстратом либо за счет изменения его нормальной функции.
• Доминантно-негативные мутации являются результатом несинонимичных мутаций или делеций/дупликаций внутри рамки считывания, но также могут быть вызваны экспансией тринуклеотидных повторов, однако такие варианты встречаются реже. Доминантно-негативные мутации — это гетерозиготные изменения, которые приводят к продукции аномального белка, влияющего на нормальное функционирование белка дикого типа.
е) Нормальная изменчивость генома. У каждого из нас в геноме есть от 5 до 50 млн вариантов, которые встречаются примерно через каждые 300 оснований. Эти варианты преимущественно представлены полиморфизмами, возникающими более чем у 1% популяции; они не влияют или незначительно влияют на функцию генов и клеток и не связаны с высоким риском заболеваний. Полиморфизмы могут возникать в экзонах, интронах или межгенных областях, которые составляют 98—99% генома человека. Каждый из классов генетических вариантов, которые обсуждаются, присутствует в геноме как частый полиморфизм. В то же время наиболее часто встречаются однонуклеотидные полиморфизмы, при которых возникает замена одного основания.
1. Полиморфизмы и частые заболевания. Защитные полиморфизмы и полиморфизмы с негативным воздействием, связанные с частыми заболеваниями, были выявлены преимущественно в полногеномных исследованиях (genome-wide association study) и являются основой для многих тестов, предназначенных непосредственно для потребителя (direct-to-consumer test). Данные тесты направлены на определение индивидуальных профилей риска распространенных заболеваний или признаков, таких как сердечно-сосудистые заболевания, сахарный диабет и даже облысение по мужскому типу! Примером таких полиморфизмов является полиморфизм гена SLC2A9, который не только отвечает за значительную долю нормальных популяционных вариантов концентрации мочевой кислоты в сыворотке крови, но также является фактором, предрасполагающим к развитию подагры у носителей «аллелей высокого риска».
Тем не менее реальная действительность такова, что точная оценка риска невозможна до тех пор, пока не будут получены более полные представления о картине генома в целом и знания обо всем наборе патогенных и защитных полиморфизмов.
2. Эволюционный отбор. Важную роль в эволюционном отборе играют генетические варианты: выгодные варианты приводят к положительному отбору за счет улучшения репродуктивного соответствия, а вариации, снижающие репродуктивное соответствие, в ходе эволюции исключаются. С учетом этой простой парадигмы возникает заманчивое предположение о том, что все частые мутации обеспечивают преимущества, а все редкие мутации являются патогенными. К сожалению, зачастую трудно классифицировать какую-либо частую мутацию как полезную, вредную или даже нейтральную. Мутации, которые полезны в молодом возрасте и таким образом улучшают репродуктивное соответствие, на более поздних этапах жизни могут быть вредными.
Возможны мутации, дающие преимущества в выживании в определенных условиях (например, в условиях голода или пандемии), которые могут оказаться вредными при более благоприятных условиях и вызывать предрасположенность к ожирению или аутоиммунным заболеваниям.