Механизмы образования биопленок на зубах (зубного налета)
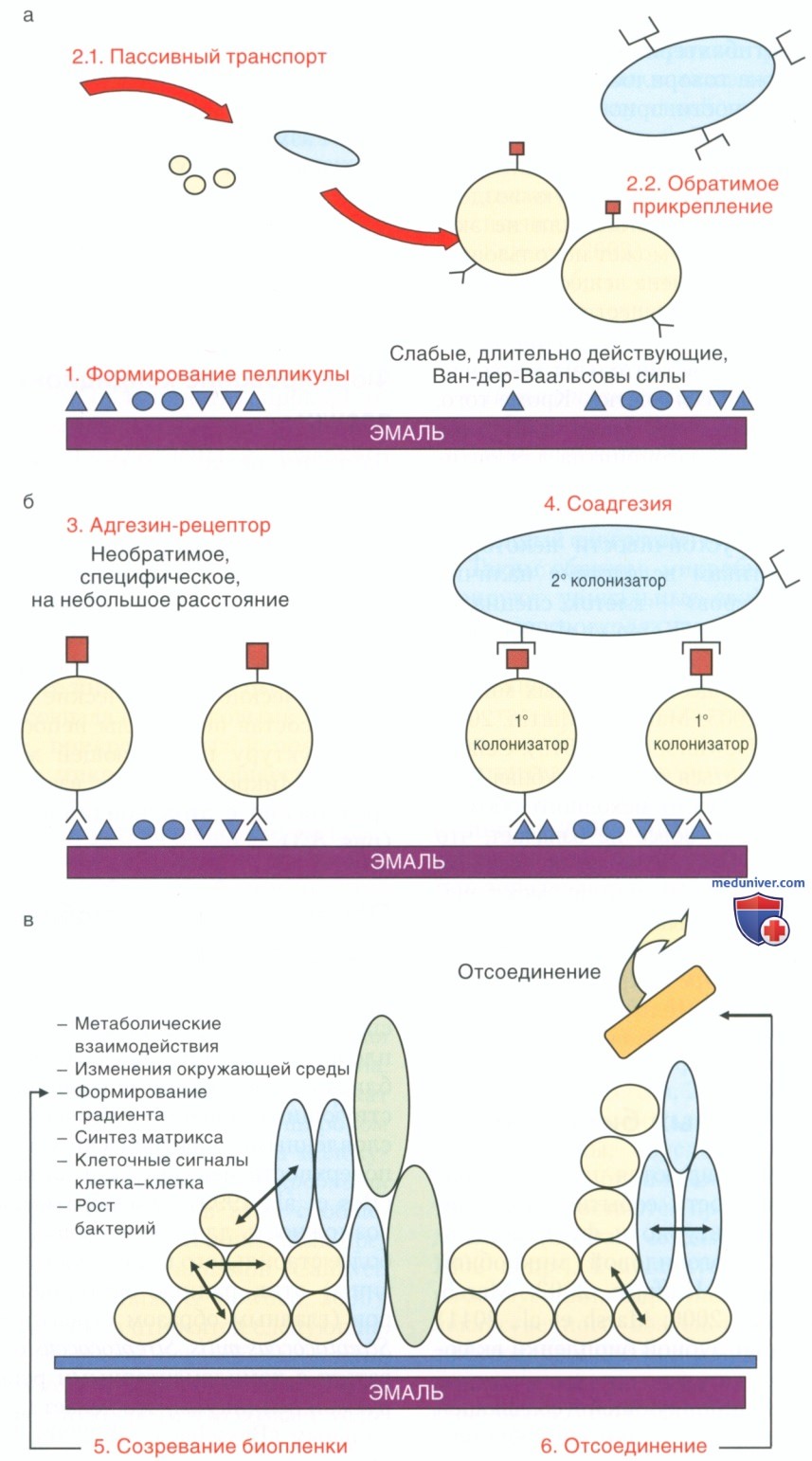
Зубные биопленки формируются путем упорядоченной последовательности событий, приводящих к образованию структурно и функционально организованной, многовидовой микробной биопленки (Socransky & Haffajee, 2002; Marsh, 2005; Kolenbrander et al., 2006; Marsh et al., 2011) (рис.2). Формирование зубной биопленки включает ряд этапов.
Рисунок 2. Схематическое представление различных этапов формирования зубных биопленок: а — формирование пелликулы на чистой поверхности зуба (1); бактерии транспортируются пассивно к поверхности зуба (2.1), где они могут обратимо удерживаться слабыми, длительно действующими силами притяжениями (2.2); б — прикрепление становится более постоянным благодаря специфическим стереохимическим молекулярным взаимодействиям между адгезинами на бактериях и комплементарными рецепторами в пелликуле (3), вторичные колонизаторы прикрепляются к уже прикрепленным первичным колонизаторам путем молекулярных взаимодействий (соадгезия) (4); в — результаты роста в созревшей биопленке, обеспечивающего широкий спектр межмикробных взаимодействий (синергических и антагонистических) (5). Иногда клетки могут отделяться для колонизации в другом месте (6).
1. Адсорбция кондиционирующей (создающей необходимые условия) пленки (приобретенная пелликула).
2. Обратимая адгезия поверхности микробных клеток к кондиционирующей пленке.
3. Более стабильное прикрепление на основе взаимодействия специфических молекул с поверхностью микробной клетки (адгезины) и комплементарными молекулами (рецепторы), присутствующими в кондиционирующей пленке.
4. Соадгезия, при которой вторичные колонизаторы прилипают к рецепторам уже присоединенных бактерий (Kolenbrander et al., 2010), что приводит к увеличению микробного разнообразия.
5. Размножение присоединенных клеток, что приводит к увеличению биомассы и синтезу экзополимеров для формирования матрикса биопленки (созревание зубного налета).
6. Отрыв прикрепленных клеток для колонизации в других местах.
Эти этапы будут теперь описаны более подробно.
а) Формирование кондиционирующей пленки. Бактерии редко заселяют чистую эмаль. Через несколько секунд после прорезывания или после очистки поверхность зубов покрывается кондиционирующей молекулярной пленкой (биологически активные белки, фосфопротеины и гликопротеины), формируемой в основном из компонентов слюны (но и из GCF, и самих бактерий) (Hannig et al., 2005).
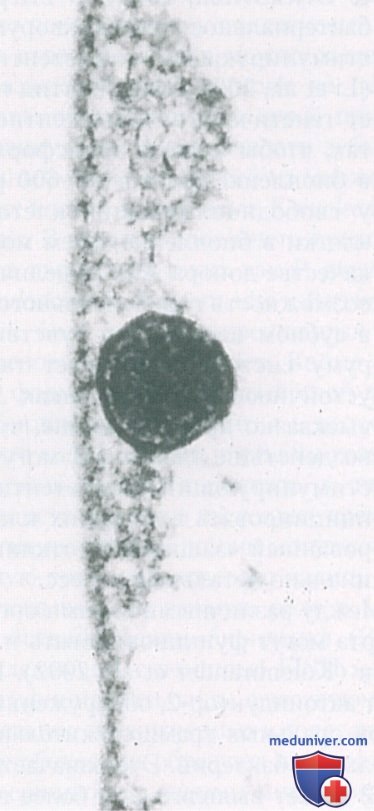
Кондиционирующая пленка изменяет биологические и химические свойства поверхности, и состав пелликулы непосредственно влияет на структуру последующей микробной колонизации. Микроорганизмы взаимодействуют непосредственно с этой кондиционирующей пленкой (рис. 3).
Рисунок 3. Электронная микрофотография, иллюстрирующая 4-часовую пелликулу с одной прикрепленной бактериальной клеткой.
б) Обратимое и более стабильное прикрепление. Первоначально только немногие виды бактерий способны прикрепляться к кондиционирующей пленке. Бактерии могут обратимо удерживаться близко к поверхности слабыми, длительно действующими физико-химическими силами, обусловленными электрическим зарядом молекул на поверхности пелликулы и на поверхности клетки (Bos et al., 1999).
Эта обратимая адгезия создает возможность для последующего более сильного и более стабильного прикрепления. Молекулы (адгезины) этих ранних бактериальных колонизаторов (главным образом стрептококков, к примеру, Streptococcus mitis, Streptococcus oralis) могут связываться с комплементарными рецепторами формируемой пелликулы, что делает прикрепление более сильным (Busscher et al., 2008; Nobbs et al., 2011).
Отдельные виды могут образовывать несколько адгезинов (Nobbs et al.); у грамположительных бактерий в качестве адгезинов могут выступать несколько семейств поверхностных белков, в том числе семейства богатых серином повторов, антигенов I/II и пилей. У грамотрицательных бактерий функции адгезинов выполняют белки-аутотранспортеры, связывающие внеклеточный матрикс, белки и пили (Nobbs et al., 2011).
в) Соадгезия. После прикрепления первичные колонизаторы начинают размножаться. Метаболизм прикрепившихся первыми бактерий изменяет локальную среду, к примеру, делая ее более анаэробной после потребления кислорода и производства восстановленных конечных продуктов обмена веществ.
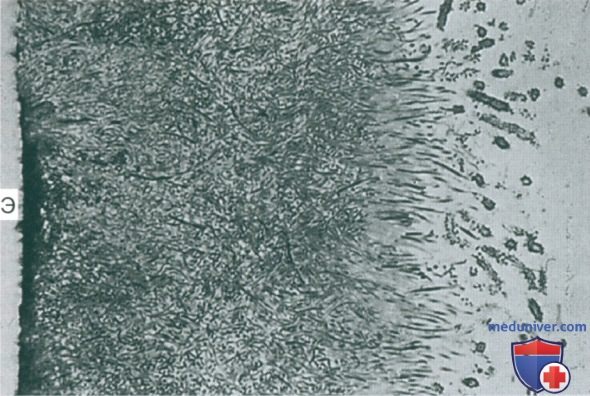
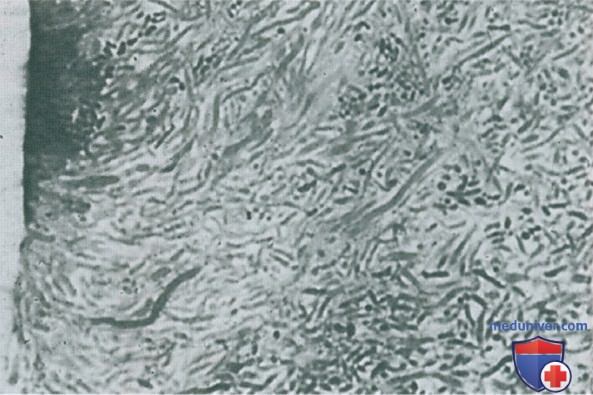
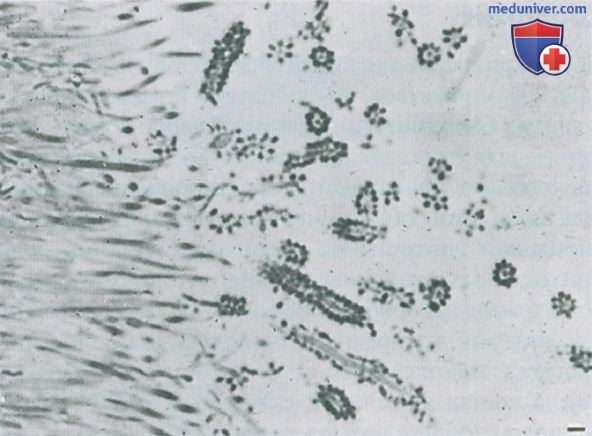
По мере развития биопленки адгезины на поверхности клеток более прихотливых вторичных колонизаторов, таких как облигатные анаэробы, связываются с рецепторами тех бактерий, которые уже прикреплены, с помощью процесса так называемой соадгезии или соагрегации, а состав биопленки становится более разнообразным (процесс называется микробной сукцессией) (Kolenbrander et al., 2006) (рис. 4, 5).
Рисунок 4. Полутонкий срез наддесневой биопленки на эмали (Э), которая была растворена до изготовления среза, х750.
Рисунок 5. Полутонкий срез наддесневой биопленки. Преобладают нитчатые бактерии. Некоторые из этих микроорганизмов на поверхности окружены кокками; конфигурация напоминает кукурузный початок. х1400.
Ключевой организм в развитии зубного налета — Fusobacterium nucleatum. Этот вид может соадгезировать большинство бактерий полости рта, а также выступать в качестве важного связующего звена между ранними и поздними колонизирующими видами. Соадгезия позволяет бактериям, находящимся вблизи других организмов, приобрести дополнительные метаболические функции.
г) Созревание налета. Некоторые из прикрепленных бактерий синтезируют внеклеточные полимеры (матрикс биопленки), которые могут упрочнять присоединение биопленки. Матрикс больше, чем просто структурная основа для биопленки; он может связывать и удерживать молекулы, в том числе ферменты, а также тормозить проникновение заряженных молекул в биопленку (Allison, 2003; Vu et al., 2009; Marsh et al., 2011).
Биопленки структурно и функционально организованы, и гетерогенные условия внутри биопленки индуцируют новые паттерны бактериальной генной экспрессии, а близкое соседство различных видов обеспечивает возможность взаимодействия (Kuramitsu et al., 2007; Hojo et al., 2009; Marsh et al., 2011). Примеры таких взаимодействий следующие.
• Формирование пищевых цепей (в которых конечный продукт метаболизма одного организма используется в качестве основного питательного вещества вторичным потребителем) и межвидового метаболического взаимодействия для усвоения структурно сложных макромолекул хозяина. Эти взаимодействия повышают метаболическую эффективность микробного сообщества (Periasamy & Kolenbrander, 2010; Marsh et al., 2011).
• Передача сигналов между клетками. Бактерии зубного налета, как было показано, взаимодействуют друг с другом зависящим от плотности клеток образом через небольшие диффундирующие молекулы. При этом они используют стратегии, аналогичные описанным для других биопленок, например секреции малых пептидов грамположительными бактериями для координации экспрессии генов среди клеток своего вида (Suntharalingam & Cvitkovitch, 2005).
У Streptococcus mutans бактериальное чувство кворума опосредуется стимулирующим компетентность пептидом (Li et al., 2002). Этот пептид также индуцирует генетическую компетентность у S. mutans так, чтобы частота трансформации S. mutans в биопленке была в 10-600 раз больше, чем у свободноплавающих клеток. Лизируемые клетки в биопленке затем могут выступать в качестве донора ДНК, увеличивая тем самым возможность горизонтального переноса генов в зубном налете.
Эта чувствительная к кворуму система регулирует также кислотную устойчивость в биопленках S. mutans. Было высказано предположение, что S. mutans при воздействии низких pH могут высвобождать стимулирующий компетентность пептид и инициировать в соседних клетках координированный «защитный» отклик на этот потенциально летальный стресс.
Между различными видами организмов в полости рта могут функционировать и другие системы связи (Kolenbrander et al., 2002). Гены LuxS кодируют автоиндуктор-2, обнаруженный у нескольких родов оральных грамположительных и грамотри-цательных бактерий. Это означает, что автоиндук-тор-2 может выявляться у более широкого диапазона видов. Несколько предполагаемых патогенов пародонта (F. nucleatum, Р. intermedia, Р. gingivalis, Aggregatibacter actinomycetemcomitans) секретируют сигнальные молекулы, связанные с автоиндуктором-2 (Fong et al., 2001; Frias et al., 2001).
Рисунок 6. На поверхности биопленки видны формации в виде кукурузного початка, показанные на рис. 4 и 5. х1300. Срез 1 мкм.
В зрелом зубном налете можно увидеть характерные клеточные ассоциации, похожие на кукурузный початок (в котором коккоподобные клетки прикрепляются к нитчатым) (рис. 6), и похожие на ершик формации (палочкоподобные бактерии прикрепляются перпендикулярно бактериальным нитям) (Zijnge et al., 2010). Они будут обсуждаться в следующем разделе.
Структура биопленок повышает вероятность успешного горизонтального переноса генов. Как уже говорилось выше, в биопленках сигнальные молекулы, такие как стимулирующий компетентность пептид, заметно повышают способность клеток-реципиентов поглощать ДНК. На модели биопленок была продемонстрирована передача у стрептококков конъюгативных транспозонов, кодирующих устойчивость к тетрациклину.
Выделение из носоглотки резидентных (S. mitis, S. oralis) и патогенных (S. pneumoniae) бактерий с генами устойчивости к пенициллину показало общую мозаичную структуру, что подтверждает перенос генов в естественных условиях (Dowson et al., 1990; Hakenbeck et al., 1998). Аналогичные данные свидетельствуют об обмене генов, ответственных за пенициллинсвязывающий белок, среди синантропных и патогенных Neisseria (Bowler et al., 1994).