Комплемент — сбалансированная система, которая играет фундаментальную роль в защите организма от действия чужеродных агентов. Система комплемента выполняет и др. функции: содействие фагоцитарному удалению погибающих клеток, молекулярного мусора и слабых или избыточных синапсов во время формирования мозга. Компоненты и рецепторы комплемента функционируют внутри отдельных клеток и могут стабилизировать в/клеточный гомеостаз.
Однако активация комплемента также может причинить вред и вызвать многие заболевания. Система комплемента состоит из нескольких частей:
(1) классический, лектиновый и альтернативный пути, которые взаимодействуют и зависят друг от друга в своей полной активности;
(2) комплекс мембранной атаки (C5b6789), образованный в результате активации любого пути;
(3) рецепторы клеточной мембраны, которые связывают компоненты или фрагменты комплемента для опосредования активности комплемента;
и (4) ряд многочисленных плазменных и мембранных регуляторных белков (табл. 1 и рис. 1).
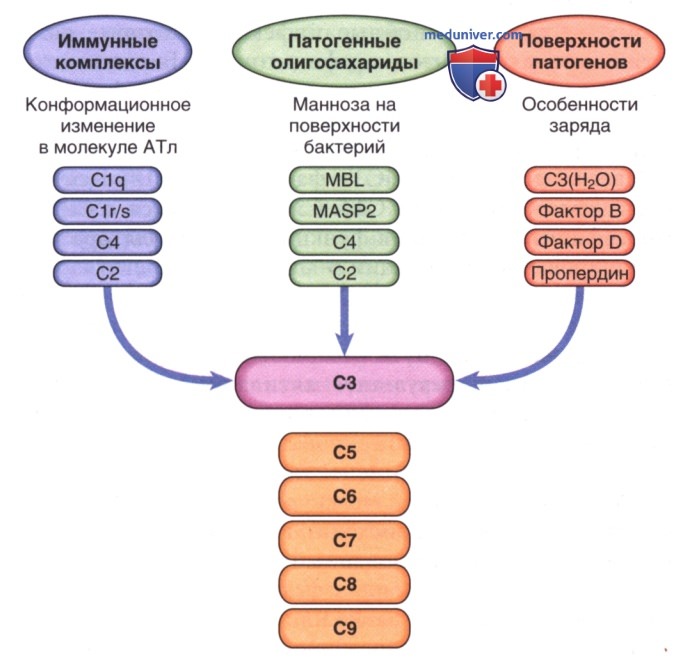
Рисунок 1. Активация каскада комплемента. Примечание. Классический путь активируется АТл, а лектин, связывающий маннозу, и альтернативные пути активируются патогенами. В каждом случае плечо активации приводит к расщеплению компонента С3.
Циркулирующие компоненты и регуляторы составляют 15% от фракции глобулинов и 4% всех белков плазмы. Нормальные уровни концентрации компонентов комплемента в сыворотке зависят от возраста. У новорожденных отмечается легкая или умеренная степень недостаточность всех компонентов комплемента.
После С1423 номенклатура комплемента логична и подчиняется всего нескольким правилам. Фрагментам компонентов, полученным в результате расщепления др. компонентами, действующими как ферменты, присваиваются строчные буквы (a, b, с, d, е). Исключение — фрагменты С2: меньшему фрагменту, который выделяется в окружающие жидкости, присваивается строчная буква «а», и большая часть молекулы, связанная с др. компонентами или с некоторой частью иммунного комплекса, обозначается буквой «b», напр., С3а и С3b. Компоненты альтернативного пути, В и D, обозначены прописными буквами, как и регуляторный I- и Н-белки, которые ингибируют оба пути. СЗ и его главный фрагмент С3b — компоненты классических и альтернативных путей.
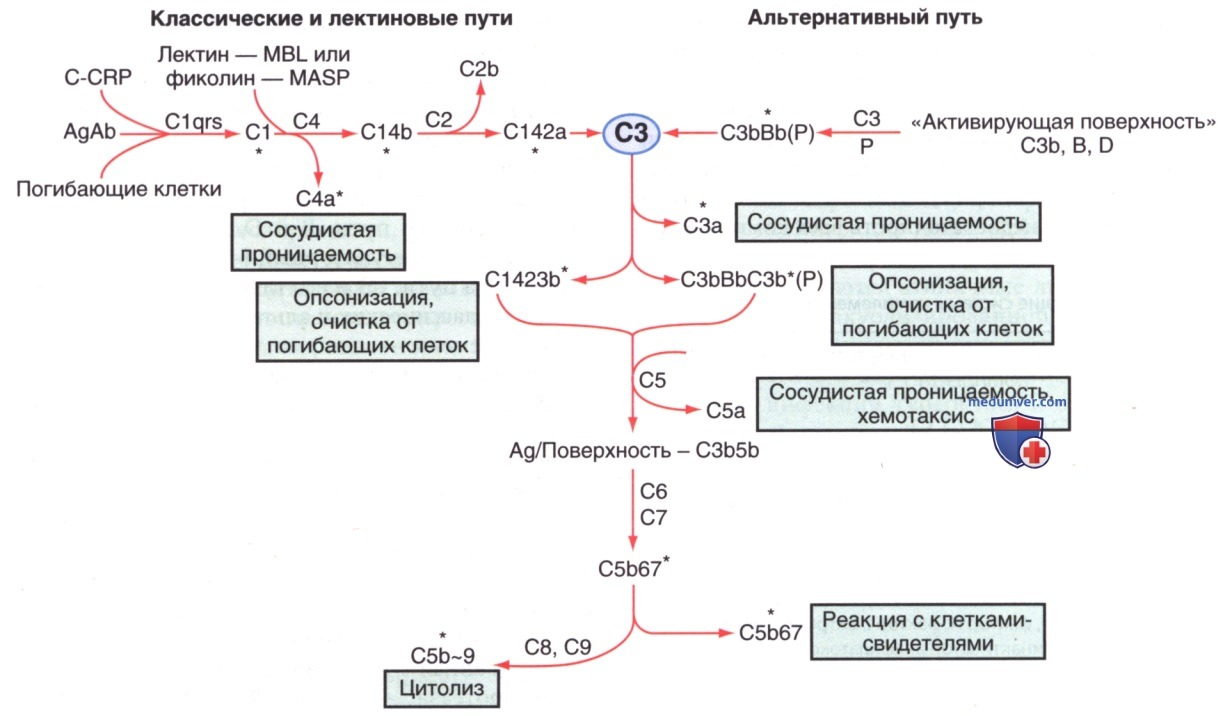
Комплемент — система взаимодействующих белков. Биологические функции системы зависят от взаимодействий отдельных компонентов, которые происходят последовательно (каскадно) одно за другим. Активация каждого компонента, кроме 1-го, зависит от активации предыдущего компонента или компонентов в последовательности. Взаимодействие происходит по трем направлениям (рис. 2): классический путь в последовательности АГн — АТл — С142356789; лектиновый путь (связывание углеводов) в последовательности микробный углевод-лектин (маннозо-связывающий лектин или фиколин) — ассоциированная с маннозо-связывающим лектином сериновая протеаза — С42356789; и альтернативный путь в последовательности активатор — C3bBD — С356789.
Рисунок 2. Каскад активации компонентов классического и пектинового путей комплемента и взаимодействие с альтернативным путем. Примечание. MBL — маннозосвязывающий лектин; MASP — MBL-ассоциированная сериновая протеаза. Основная цель — активация СЗ. Функциональная деятельность, возникшая в результате активации, указывается в прямоугольных рамках. Множественные очаги, в которых проявляют активность белки-ингибиторы и белки-регуляторы (не показаны) обозначены звездочками, чтобы подчеркнуть существующий баланс между действием и регуляцией в пределах этой системы, что необходимо для защиты хозяина, но способно причинить серьезный ущерб его тканям. Ab — АТл (lg класса G или М); Ад — АГн (бактерия, вирус, опухоль или тканевая клетка); В, D, I, Р — факторы В, D, I и пропердин; C-CRP — углеводно-углеводно-реактивный белок; С4-bp — С4-связывающий белок.
АТл ускоряет скорость активации альтернативного пути, но активация может происходить на соответствующих поверхностях в отсутствие АТл. Классический и альтернативный пути взаимодействуют друг с другом благодаря обоюдной способности активировать СЗ.
Активация компонентов комплемента раннего действия (С1423) приводит к генерации активных ферментов C1, С42 и С423 на поверхности иммунного комплекса или основной клетки. Эти ферменты расщепляют и активируют следующий компонент в последовательности. Взаимодействие между С5b, С6, С7, С8 и С9 неферментативно и зависит от изменений молекулярной конфигурации.
а) Классические и лектиновые пути. Каскад классического пути начинается с фиксации С1 посредством Clq к Fc, не-АГн-связывающей части молекулы АТл после взаимодействия АГн-АТл. Трикомплекс С1 меняет конфигурацию, и подкомпонент C1s становится активным ферментом С1 эстеразой. Некоторые бактерии, РНК-вирусы и липидный компонент бактериального эндотоксина могут непосредственно активировать C1q, запуская каскад полного комплемента.
В пределах врожденного иммунного ответа реактивные «естественные» АТл и СРВ, который реагирует с углеводом из микроорганизмов и с погибающими клетками, могут заменять специфическое АТл при фиксации C1q и инициировать реакцию всей последовательности. Эндогенные агенты, включая кристаллы мочевой кислоты, отложения амилоида, ДНК и компоненты поврежденных клеток, такие как апоптотические пузырьки и митохондриальные мембраны, могут активировать C1q.
Но в этом случае комплекс лиганд-C1q активно взаимодействует с ингибиторами С4-связывающего белка и фактора Н, обуславливая С3-опосредованную опсонизацию и фагоцитоз, но ограничивая системный воспалительный ответ, вызываемый микробами. Белок C1q, синтезируемый клетками ГМ и сетчатки, прикрепляется к избыточным синапсам, которые затем очищаются посредством рецепторов C1q в микроглии, освобождая путь новым синапсам для заселения развивающейся НС.
Четыре узнающие молекулы лектинового пути: маннозо-связывающий лектин и фиколины 1, 2 и 3. Маннозосвязывающий лектин — белок семейства углеводсвязывающих белков (лектинов), которые играют важную роль во врожденном неспецифическом иммунитете. Его структура гомологична структуре Clq. Эти лектины в сочетании с ассоциированными с маннозо-связывающим лектином сериновыми протеазами 1, 2 и 3 (MASP 1, 2, 3) могут связываться с маннозой, липотейхоевой кислотой и др. углеводами на поверхности бактерий, грибов, паразитов и вирусов. Затем MASP функционируют как С1, расщепляя С4 и С2 и активируя каскад комплемента. Пептид С4а обладает слабой «анафилатоксиновой» активностью и реагирует с тучными клетками, высвобождая химические медиаторы немедленной гиперчувствительности, включая гистамин.
Высвобождаемые вслед за ними С3а и С5а — мощные анафилатоксины, а С5а — важный хемотаксический фактор. Фиксация С4b к комплексу обуславливает его соединение с нейтрофилами, макрофагами, В-клетками, дендритными клетками и эритроцитами. MASP-2 может активировать свертывание, трансформируя протромбин в тромбин, что предотвращает распространение микробов.
Следующий этап — расщепление С3 и образование С3b. Плазменная концентрация С3 наиболее высокая по сравнению с концентрациями остальных компонентов, поэтому его активация — наиболее важный этап с точки зрения биологической активности. Расщепление С3 происходит посредством СЗ-конвертазы классического пути, С142, или посредством СЗ-конвертазы альтернативного пути (С3bВb). С момента прикрепления С3b к мертвой или умирающей клетке хозяина он может связываться с клетками, содержащими рецепторы для С3b (рецептор комплемента 1 — CR1), включая В-лимфоциты, эритроциты и фагоцитарные клетки (нейтрофилы, моноциты и макрофаги). Для осуществления эффективного фагоцитоза большинству микроорганизмов, особенно нейтрофилам, необходимо связывание С3 с микробом.
Подтверждение этому — тяжелые гнойные инфекции, которые часто возникают у пациентов с дефицитом С3. Биологическая активность С3b регулируется расщеплением фактора I до iC3b, что способствует фагоцитозу при связывании с рецептором iC3b (CR3) на фагоцитах. В результате дальнейшего расщепления iC3b фактором I и протеазами образуется C3dg, затем C3d. C3d связывается с CR2 на В-лимфоцитах, выполняя функцию ко-стимулятора АГн-индуцированной активации В-клеток.
б) Альтернативный путь. Альтернативный путь может быть активирован С3b, генерируемым в результате активации классического пути, протеазами нейтрофилов или системой свертывания. Он также может активироваться формой С3, образованной в результате наименее интенсивной, спонтанной реакции нативного С3 с молекулой воды — «глушение», которое постоянно происходит в плазме. После образования С3b или гидролизованный С3 может связываться с любой соседней клеткой или с фактором В. Фактор В, присоединенный к С3b в плазме или на поверхности, может расщепляться до Вb сывороточной протеазой — фактором D. Комплекс С3bВb становится эффективной конвертазой С3, которая способна к генерации большего количества С3b посредством петли амплификации. Пропердин может связываться с С3bВb, повышая стабильность фермента и защищая его от инактивации факторами I и Н, модулирующих петлю и путь.
Активации альтернативного пути способствуют «активирующие поверхности», фиксирующие С3b, включая бактериальную тейхоевую кислоту и эндотоксин, инфицированные вирусом клетки, комплексы АГн-IgA, системы искусственного кровообращения, а также мембраны для почечного диализа. Эти поверхности действуют, защищая фермент С3bBb от регулирования, которое осуществлялось бы факторами I и Н. Мембрана кроличьих красных кровяных телец — поверхность, которая служит основой для анализа активности альтернативного пути с использованием сыворотки крови. Сиаловая кислота на поверхности микроорганизмов или клеток предотвращает образование эффективного альтернативного пути С3-конвертазы, стимулируя активность факторов I и Н. Значительная активация С3 может происходить по альтернативному пути, а результирующая биологическая активность качественно не отличается от таковой при активации С142 (см. рис. 2).
в) Мембранный атакующий комплекс. Цепочка преобразований, ведущая к цитолизу, начинается с прикрепления С5b к С5-активирующему ферменту классическим (C4b2a3b) или альтернативным путем (С3bВb3b). С6 связывается с С5Ь без этапа расщепления, стабилизируя активированный фрагмент С5b. Затем комплекс С5b6 диссоциирует от С423 и взаимодействует с С7. Комплексы С5b67 должны быстро присоединяться к мембране родительской клетки или клетки-свидетеля: в противном случае они теряют свою активность. Затем С8 взаимодействует с комплексом C5b678, что способствует добавлению множественных молекул С9. Полимер С9, состоящий из 3-6 молекул, образует трансмембранный канал, и происходит лизис.
г) Механизмы регуляции. Без механизмов регуляции, действующих в нескольких точках, не было бы эффективно работающей системы комплемента, а неконтролируемое потребление компонентов нанесло бы серьезный и даже смертельный вред больному. На первом этапе ингибитор C1 (С1 INH) подавляет ферментативную активность С1r и C1s. Расщепление С4 и С2. C1 INH также ингибирует MASP-2, факторы XIа и XIIа системы свертывания крови и калликреин контактной системы. Активированный С2 характеризуется коротким периодом полураспада, и эта относительная нестабильность ограничивает продолжительность эффективности С42 и С423. Активирующий С3 фермент альтернативного пути, С3bВb, также характеризуется коротким периодом полураспада, который продлевается за счет связывания пропердина (фактор Р) ферментным комплексом. Пропердин также может напрямую связываться с микробами и способствовать сборке конвертазы СЗ альтернативного пути.
Плазма содержит фермент карбоксипептидазу N, который отщепляет аргинин на N-конце белка от С4а, С3а и С5а, тем самым ограничивая их биологическую активность. Фактор I инактивирует С4b и С3b; фактор Н ускоряет инактивацию С3b фактором I; и аналогичный фактор, С4-связывающий белок (С4-bp), ускоряет расщепление С4b фактором I, тем самым ограничивая сборку С3 конвертазы. Три белковых компонента клеточных мембран — CR1, мембранный белок-кофактор (МСР) и фактор ускорения распада (DAF) — способствуют разрушению С3 и С5 конвертаз, собранных на этих мембранах. Другой белок, ассоциированный с клеточной мембраной, CD59, может связывать С8 или С8 и С9, тем самым препятствуя встраиванию комплекса мембранной атаки (С5b6789).
Белки плазмы крови витронектин и класте-рин могут ингибировать прикрепление комплекса С5b67 к клеточным мембранам, связывать С8 или С9 в полный комплекс мембранной атаки, или иным образом препятствовать образованию или включению этого комплекса. Витронектин также способствует поглощению макрофагами умирающих нейтрофилов. Гены регуляторных белков фактора Н, С4-bр, МСР, DAF, CR1 и CR2 сгруппированы на хромосоме 1.
д) Участие в защите организма больного. Нейтрализация вируса АТл может быть усилена с помощью С1 и С4 и дополнительно усилена фиксацией С3b классическим или альтернативным путем. Именно поэтому комплемент особенно важен на ранних стадиях вирусной инфекции при недостаточном количестве АТл. АТл и каскад компонентов комплемента также устраняют вирулентность микроорганизмов за счет образования характерных «дырок» комплемента, что видно на электронной микроскопии. Фиксация C1q может опсонизировать (способствовать фагоцитозу) через связывание с рецептором C1q фагоцита.
С4а, С3а и С5а могут связываться с тучными клетками и тем самым запускать высвобождение гистамина и др. медиаторов, что приводит к расширению сосудов, отеку и воспалительной гиперемии. С5а может усиливать фагоцитоз С3b-опсонизированных макрофагами частиц и стимулировать макрофаги высвобождать цитокины, ФНО и IL-1. С5а — основной хемотаксический фактор нейтрофилов, моноцитов и эозинофилов, который может эффективно фагоцитировать опсонизированные СЗЬ или расщепленные С3b (iC3b) микроорганизмы. Дальнейшая инактивация связанного с клеткой С3b путем расщепления на C3d и C3dg устраняет его опсонизирующую активность, но он все еще может связываться с В-клетками. Фиксация С3b к клетке-мишени может усилить его лизис естественными киллерами или макрофагами.
Нерастворимые иммунные комплексы могут стать растворимыми после связывания с С3b, т.к. С3b нарушает упорядоченную решетку АГн-АТл. Связывание С3b с комплексом также позволяет ему прикрепляться к рецепторам С3 (CR1) на эритроцитах, которые затем транспортируют комплексы к макрофагам печени и селезенки для удаления. Этот феномен частично объясняет заболевание иммунных комплексов, обнаруживаемое у пациентов с отсутствием С1, С4, С2 или С3.
Система комплемента служит для связи врожденной и адаптивной иммунных систем. С4b или С3b, связанные с иммунными комплексами, способствуют их взаимодействию с АГн-презентирующими макрофагами, дендритными клетками и В-клетками. Соединение АГн с C3d позволяет связываться с CR2 на В-клетках, что значительно уменьшает количество АГн, необходимое для запуска АТл-ответа.
Нейтрализация эндотоксина in vitro и защита от его летального воздействия на экспериментальных животных требует наличия C1 INH и компонентов комплемента более позднего действия (С6). Активация всей последовательности комплемента может привести к лизису инфицированных вирусом клеток, опухолевых клеток и большинства микроорганизмов. Бактерицидная активность комплемента не важна для защиты организма больного, за исключением случаев менингококковых инфекций у пациентов с отсутствием компонентов комплемента более позднего действия.
Видео №1: схема классического пути активации комплемента
Видео №2: схема альтернативного пути активации комплемента