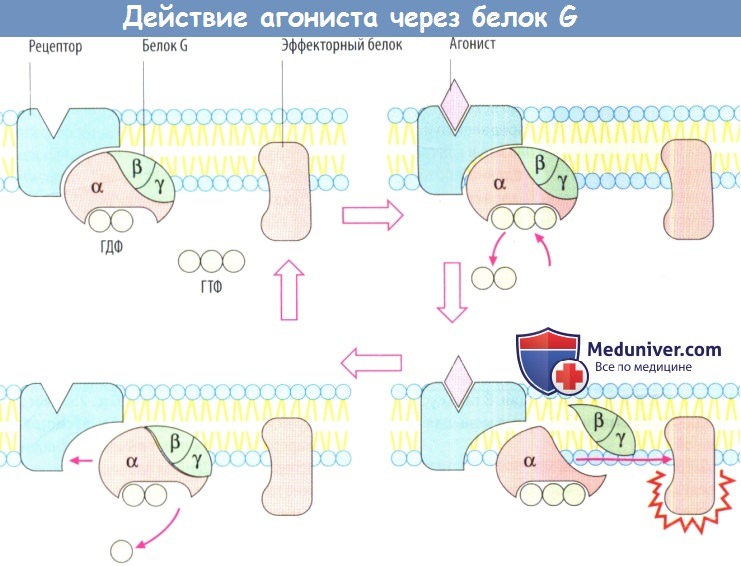
При передаче сигнала на рецепторы, сопряженные с белком G, участвует фактически такой же основной механизм. Связывание агониста рецептором приводит к изменению конформации белка рецептора. Это изменение передается на белок G: а-субъединица изменяет гуанозиндифосфат (ГДФ) на гуанозинтрифосфат (ГТФ), затем диссоциирует от двух других субъединиц, связывается с эффекторным белком и изменяет его функциональное состояние.
В принципе, β- и γ-субъединицы тоже способны взаимодействовать с эффекторными белками, а-субъединица медленно гидролизует связанный ГТФ до ГДФ. Ga-ГДФ не обладает аффинитетом к эффекторному белку и вновь связывается с β- и γ-субъединицами. Белки G могут диффундировать вдоль мембраны; они не прикреплены к отдельным белкам рецепторов. Тем не менее существует связь между типами рецепторов и типами белка G.
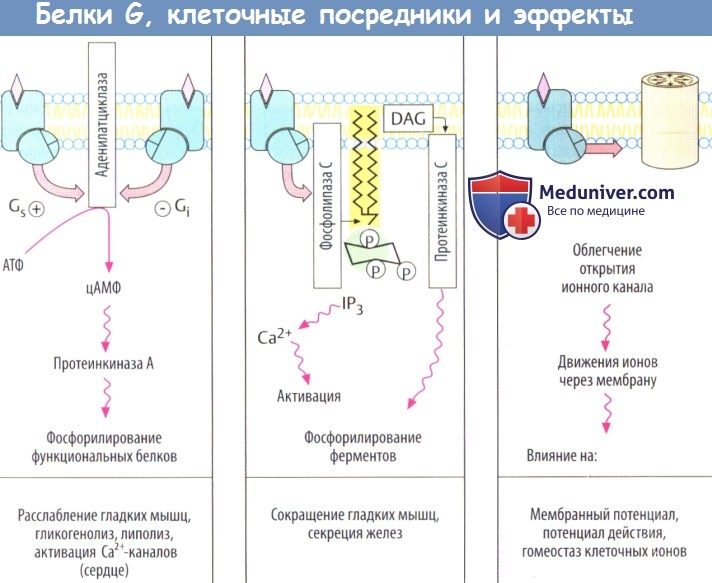
Более того, α-субъединицы отдельных белков G различаются аффинитетом к разным эффекторным белкам, а также характером воздействия на них. Ga-ГТФ белка Gs стимулирует аденилатциклазу, в то время как Ga-ГТФ белка Gi является ее ингибитором.
Группа сопряженных с G-белком рецепторов включает мускариновые холинорецепторы, адренорецепторы норадреналина и адреналина, а также рецепторы дофамина, гистамина, серотонина, глутамата, ГАМК, морфина, простагландинов, лейкотриенов и многих других медиаторов и гормонов.
К основным эффекторным белкам у сопряженных с G-белком рецепторов относятся аденилатциклаза (АТФ => внутриклеточный посредник цАМФ), фосфолипаза С (фосфатидилинозитол => внутриклеточные посредники инозитолтрифосфата и диацилглицерола), а также белки ионного канала. Множество клеточных функций регулируется концентрацией клеточного циклического аденозинмонофосфата (цАМФ), поскольку цАМФ увеличивает активность протеинкиназы А, которая катализирует превращение фосфатных групп в функциональные белки.
Повышение уровня цАМФ приводит к снижению тонуса гладких мышц, увеличению сократимости сердечной мышцы, а также повышению гликогенолиза и липолиза. Фосфорилирование белков сердечных Са2+-каналов повышает вероятность открытия каналов во время деполяризации мембраны. Следует отметить, что цАМФ инактивируется фосфодиэстеразой. Ингибиторы данного фермента повышают внутриклеточную концентрацию цАМФ и действуют как адреналин.
Белок рецептора самостоятельно фосфорилируется с потерей свойства активировать связанный белок G. Это один из механизмов, который способствует снижению чувствительность клетки во время длительной стимуляции рецептора агонистом (десенситизация).
Активация фосфолипазы С приводит к гидролизу мембранного фосфолипида фосфатидилинозитол-4,5-дифосфата до инозитолтрифосфата (IР3) и диацил-глицерола (DAG). Инозитолтрифосфат стимулирует высвобождение Са2+ из органелл-депо, что приводит к сокращению гладкомышечных клеток, распаду гликогена или к активации экзоцитоза. Диацилглицерол стимулирует протеинкиназу С, которая фосфорилирует ряд ферментов, содержащих серин или треонин.
Некоторые белки G вызывают открытие белков каналов. Таким образом активируются К+-каналы (действие АХ на синусовый узел, действие опиоидов на передачу нервного импульса).