Морбилливирусы характеризуются высокой степенью гомологии одноименных белков у разных представителей рода. Наиболее выраженное антигенное сходство морбилливирусов связано с белком F. Высокая степень гомологии выявлена в N-концевых участках полипептида F1 вирусов чумы плотоядных и кори, чего не обнаружено при сравнении с пятью представителями рода парамиксовирусов. Степень гомологии белка F1 вирусов чумы крупного рогатого скота и кори составляет 78,9%. Белок F вируса чумы крупного рогатого скота содержит область, состоящую из основных аминокислот (104-108), которая может подвергаться протеолизу с образованием F1 и F2. По данным других авторов, гомология аминокислотных последовательностей белка F вирусов чумы крупного рогатого скота и кори составляет 81,3%, а вирусов чумы крупного рогатого скота и плотоядных — 68,2%.
Высокой степенью гомологии характеризовались М-белки морбилливирусов. У вирусов чумы крупного рогатого скота и кори она составила 77,6%, а у вирусов чумы крупного рогатого скота и плотоядных — 78,2%. Обнаружено антигенное родство Н- и особенно NP-белков вирусов чумы крупного рогатого скота, кори и чумы плотоядных.
Белок L вируса кори имеет значительное структурное сходство со своими аналогами в вирусах Сендай и ньюкаслской болезни. Подобную закономерность обнаружили при сравнении белков М вирусов ПГ-3 человека, Сендай, кори и чумы плотоядных. Однако в структуре белков М представителей рода парамиксовирусов гомологии не обнаружено. Моноклональные антитела к белку F-вируса кори нейтрализовали инфекционность и фузогенную активность вируса, а также защищали мышей при инфицировании гомологичным вирусом. Введение крысам гликопротеинов Н и F вируса кори обеспечивало такой же иммунитет, как и введение цельновирусного препарата. Эти данные свидетельствуют о важной роли обоих гликопротеинов в создании иммунитета при морбилливирусных инфекциях, для возбудителей которых характерно тесное структурное и молекулярно-биологическое сходство. Одной из особенностей вируса кори является способность его к длительной персистенции в нейронах и глиальных клетках мозга реконвалесцентов. Это явление связывают с гипермутацией вируса, которая, как полагают, является одноступенчатым процессом.
Вирус мелких жвачных (овец и коз) не является вариантом вируса чумы крупного рогатого скота и по степени родства в пределах рода морбилливирусов далек от него так же, как от вируса кори. Морбилливирусы, вызывающие массовое заболевание, а также гибель тюленей, дельфинов и морских свинок, оказались идентичными или близкородственными вирусу чумы плотоядных.
Среди представителей рода пневмовирусов особое место занимают респираторно-синцитиальные (PC) вирусы, вызывающие широко распространенные поражения нижних дыхательных путей у детей, телят, овец и коз.
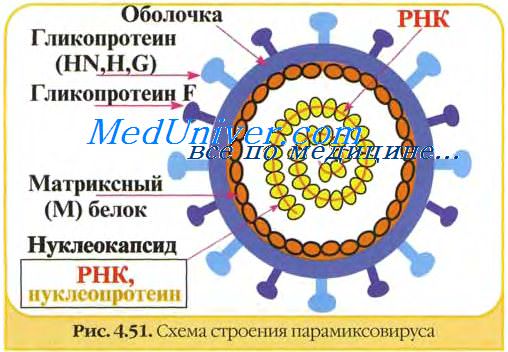
PC-вирус отличается от других парамиксовирусов большим количеством идентифицированных структурных белков, порядком генов, отсутствием гемагглютинина и нейраминидазы, а также диаметром нуклеокапсида (12—13 нм) и длиной поверхностных шипиков( 0— 12нм). PC-вирусы имеют 10 генов, которые транскрибируются с образованием 10 мРНК [940]. В структуре PC-вирусов идентифицировано 9 или 10 полипептидов, два из которых G и F гликозированны и располагаются на поверхности вириона. Белок F состоит из двух субъединиц и выполняет функцию слияния, белок G — прикрепительную функцию.
В белке слияния F PC-вируса крупного рогатого скота один из двух антигенных участков ответственен за нейтрализацию. Гликопротеины G и F оказались ответственными за индукцию ВН-антител и образование протективного иммунитета. МАТ к белкам оболочки вириона нейтрализовали инфекционную и синцитийобразующую активность PC-вируса крупного рогатого скота и выявляли (МФА) оболочечные гены вируса, эксперессированные на поверхности инфицированных клеток, а также вызывали их лизис в присутствии комплемента.
Неструктурный белок NS2 PC-вируса крупного рогатого скота не существенен для его репликации in vitro, но его присутствие способствует этому. Мутант, лишенный гена NS2, не реверсирует и представляет интерес в качестве вакцинного штамма.
Мыши, иммунизированные белками F и G PC-вируса, приобретали полную устойчивость к суперинфицированию гомологичным вирусом. Иммунизация белками N и М2 создавала частичную защиту, тогда как другие белки (HN, P, SH, М, 1В или 1С) не обладали протективными свойствами. При сравнении белков РС-вирусров человека и крупного рогатого скота основные антигенные различия выявлены в гликопротеине G. Белки слияния этих вирусов гомологичны на 83-84%. Наибольшая дивергенция связана с субъединицей F2 в области, предшествующей протеолитическому расщеплению белка F. По белкам N гомология составила 93,3%.
Выявлены две подгруппы А и В PC-вирусов человека и крупного рогатого скота, различающиеся между собой структурой белков. Более тесным антигенным родством характеризовались вирусы подгруппы А. Поскольку прикрепительный гликопротеин G обеспечивал протективный эффект только против гомологичной подгруппы PC-вирусов, для обеспечения надежной защиты требовалось ввести в вакцинный препарат G белок обеих (А и В) антигенных субгрупп. Сыворотки крови морских свинок, привитых вакцинами из субединиц PC-вирусов человека и крупного рогатого скота, обладали хорошей перекрестной реактивностью и нейтрализовала PC-вирусы как человека, так и крупного рогатого скота.
Кроме антител, в защите от PC инфекции важная роль принадлежит Т-клеточному иммунитету. Идентифицированы эпитопы, индуцирующие Т- и В-клеточные реакции иммунитета при респираторно синцитиальной инфекции. С-концевой фрагмент белка 1А содержит два участка, стимулирующих Т-лимфоциты. Белки F и G при разных способах введения вызывали образование локальных и системных ВН-антител, а также антителозависимых цитотоксических Т-лимфоцитов.
Преставитель рода метапневмовируса — вирус ринотрахеита индеек — оказался антигеннородственным с вирусом пневмонии мышей. Значительная гомология белков HN и F у некоторых представителей парамиксовирусов — хорошая предпосылка для их использования в качестве гетерологичных живых вакцин.
Для размножения парамиксовирусов используют клеточные культуры различного происхождения. Для размножения морбилливирусов и пневмовирусов обычно ис пользуют культуры клеток гомологичных видов, но и при этом они размножают ся с трудом даже после адаптации. Размножение вирусов обычно сопровождается лизисом клеток. Образование синцития наблюдают как в культуре, так и в организме с образованием ацидофильных включений в цитоплазме. Морбилливирусы вызывают образование ацидофильных включений в ядре. Вирусы парагриппа и некоторые морбилливирусы (но не пневмовирусы) вызывают гемадсорбцию.