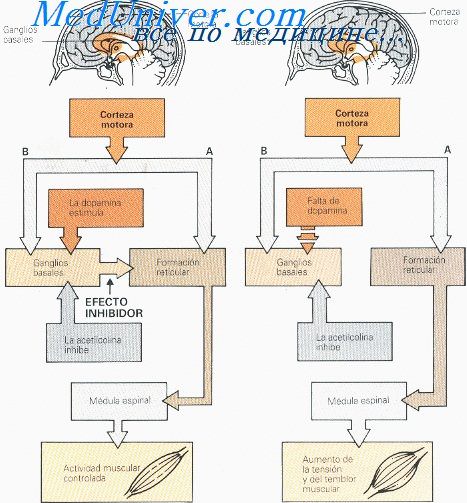
Причины болезни Паркинсона. Формы семейной болезни Паркинсона
Нами было проведено прямое секвенирование гена а-синуклеина в 9 российских семьях с несомненным аутосомно-доминантным наследованием болезни Паркинсона, при этом мутаций в кодирующей области а-синуклеина выявлено не было. Результаты исследований различных авторов и наши собственные данные свидетельствуют о том, что мутации гена а-синуклеина представляют собой весьма редкую причину семейных форм болезни Паркинсона [Vaughan J. et al., 1998; Scott W. et al, 1999].
Следующий ген аутосомно-доминантной болезни Паркинсона был выделен в 1998 году. Он локализован на хромосоме 4р14 (локус PARK5) и кодирует синтез убиквитин-карбокси-терминальной гидролазы L1 (UCH-L1) [Leroy E. et al., 1998]. Данный белок составляет 1-2% от всего белкового состава мозга, обнаруживается (как и а-синуклеин) в тельцах Леви и играет важную роль в процессах протеолиза полимерного убиквитина. Предварительные данные свидетельствуют о том, что частота мутаций в гене UCI1-L1 среди всех семейных случаев болезни Паркинсона чрезвычайно низка [Lincoln S. et al., 1999], однако в целом генетика данной молекулярной формы болезни Паркинсона остается малоизученной.
Наконец, еще в одной обширной родословной М. Farrer с соавторами в 1999 году обнаружили сцепление локуса болезни Паркинсона (PARK4) с хромосомной областью 4р14—16.3; в данной семье (как отмечалось в разделе 3.2.4.1) «мутантный» гаплотип был ассоциирован не только с болезнью Паркинсона, но также со случаями «чистого» эссенциального тремора [Farrer M. et al., 1999]. Указанное наблюдение может свидетельствовать об определенной этиологической взаимосвязи эссенциального тремора и, как минимум, одной из генетических форм болезни Паркинсона.
В литературе описаны также семьи с аутосомно-доминантной болезнью Паркинсона, в которых все вышеперечисленные генетические локусы были исключены [Gwinn-HardyK. et al., 2000].
Таким образом, уже сейчас вполне очевидно многообразие генетических вариантов семейного идиопатического паркинсонизма. По-видимому, развитие заболевания может быть обусловлено повреждением большого числа генов и их белковых продуктов, взаимосвязанных между собой посредством участия в функционировании общих метаболических путей (например, регуляции процессов протеолиза, обеспечении нейронального белкового транспорта и т.п.).
С этих позиций формирование амилоидоподобных белковых комплексов, входящих в состав телец Леви, может представлять собой универсальную патологическую реакцию как результат нарушенного на различных этапах процессинга нейрональных белков [Spacey S., Wood N., 1999].
Генетическая гетерогенность семейных случаев болезни Паркинсона существенно осложняет проведение ДНК-диапюстики в отягощенных семьях. Тотальный скрининг кодирующей области генов а-синуклеина и UCII-L1 является с практической точки зрения нецелесообразным ввиду чрезвычайной редкости мутаций в них при семейном паркинсонизме. То же самое, по-видимому, будет относиться и к каждому другому конкретному гену, когда они станут доступными для прямого анализа. Одним из путей повысить эффективность ДНК-тестирования является выделение отдельных подгрупп больных, в каждой из которых особенности клиники могли бы помочь правильному выбору генов для мутационного скрининга.
Например, для мутаций в локусе PARK1 (ген а-синуклеипа) характерно более раннее начало и быстрое прогрессирование болезни [Polymeropoulos M. el al., 1997], для РАRК4-ассоциированной формы (как уже было отмечено) характерно сочетание в семье случаев болезни Паркинсона и эссенциального тремора [Fairer M. et al., 1999] и т.д. Сейчас мы находимся лишь на самом начальном этапе анализа клинико-генетических корреляций и определения характерного фенотипа отдельных генетических форм болезни Паркинсона. Дальнейшее накопление и обобщение этих данных на основе широкого международного сотрудничества позволит выработать более четкий алгоритм для проведения ДНК-диагностики и медико-генетического консультирования в семьях, отягощенных наследственными случаями болезни Паркинсона.