Функции и физиология нижних бугорков четверохолмия (НБЧ)
а) Классификация обусловленной звуком активности центрального ядра. Физиологические свойства центрального ядра в значительной степени изменяются под воздействием анестезии барбитуратами. Этих влияний можно избежать, записывая активность отдельных единиц без анестезии на препаратах, подвергнутых децеребрации. Способность начинать движения или испытывать боль исключается путем пересечения всех волокон, приходящих и отходящих от коры головного мозга на уровне таламуса. Ограничением такого подхода является то, что нисходящая обратная связь с корой головного мозга также исключается из нейронной сети слухового анализатора в среднем мозге.
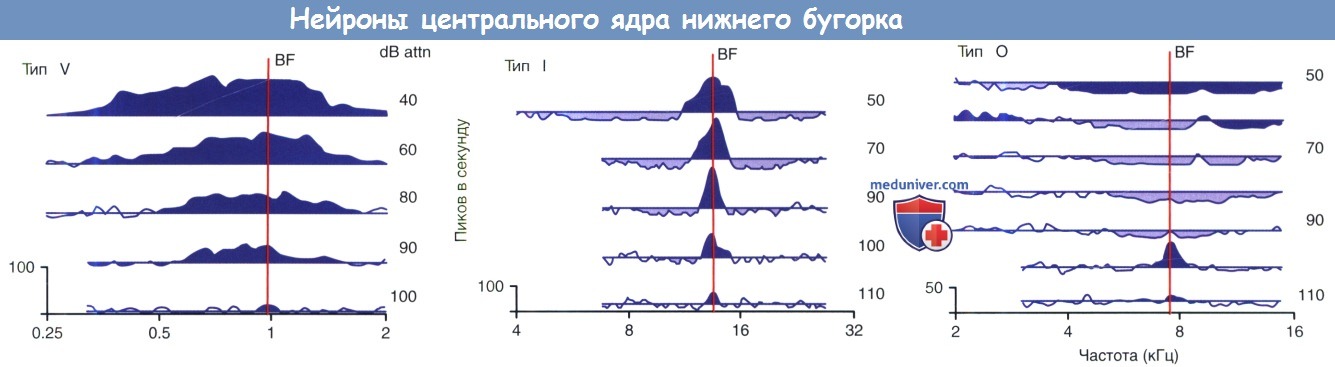
Основные клетки нижних бугорков четверохолмия (НБЧ) демонстрируют три основных варианта ответа при записи карт частотных ответов (КЧО) у децеребрированных кошек. Наиболее часто отмечался ответ, обозначенный как единица типа О, так как возбудимое рецепторное поле нейрона ограничивается небольшим О-образным островком частот около порога. В остальной части рецепторного поля доминирует подавление. Нейроны также имеют склонность к нечувствительности к сигналам от РПС. Такие особенности ответа напоминают монауральные проекционные нейроны в ДКЯ (4-ый тип нейронов). Единицы типа-О заглушаются, когда проведение блокируется в ДАП, все сильнее поддерживая прямую связь между ДКЯ и типом ответа. Единицы типа-О реже встречаются у грызунов (морские свинки, мыши, песчанки), что косвенно может указывать на их особую роль в хищных повадках кошки. Точность локализации звука у кошки значительно превышает таковую у грызунов, на которых она охотится.
Единицы типа-1 характеризуются узким I-образным рецепторным полем, которое окружено латеральным подавлением. Нейроны имеют тенденцию к настройке на высокие частоты и демонстрируют сквозной тип РПС чувствительности. Такой тип физиологических характеристик предполагает, что единицы типа I являются целью среднего мозга для восходящих возбуждающих проекций от контралатерального ЛВО. Предполагается, что контролирующее влияние этих нейронов исходит из ипсилатерального МВО.
Схема классификации по КЧО полезна в интерпретации функциональных связей нижних бугорков четверохолмия (НБЧ). Эта простая концептуальная структура показывает, как физиологически детерминированные проводящие пути от улиткового ядра остаются изолированными на пути к более высоким уровням обработки. Она также представляет меру для оценки того, насколько местные трансформации влияют на качество кодирования информации. В то же время анатомические данные ясно свидетельствуют о том, что центральное ядро является ключевым звеном слухового анализатора; то есть местом, где разрозненные репрезентации из множества входящих источников связываются воедино, формируя опыт восприятия.
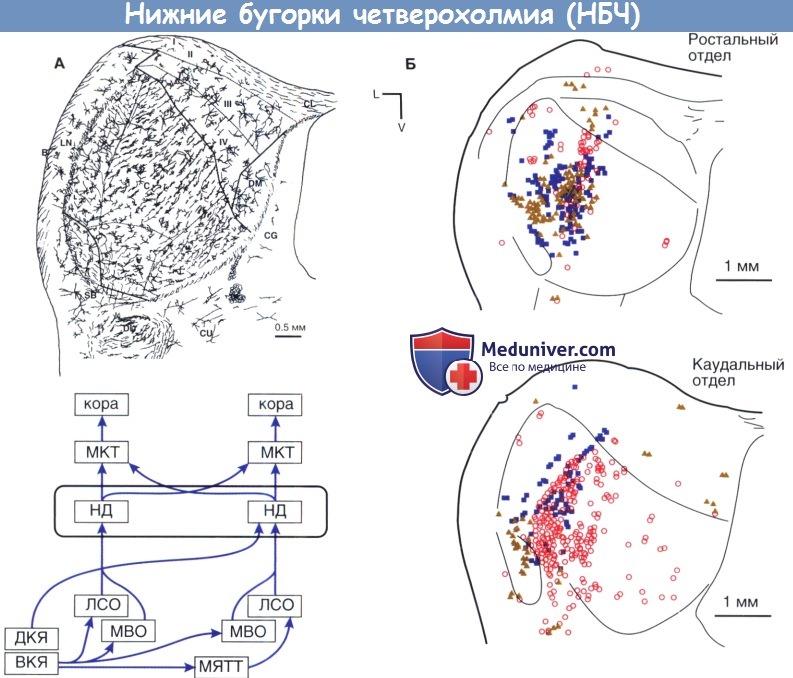
Анатомическая организация нижних бугорков четверохолмия (НБЧ) кошки.
А. Ламинарная структура, на которую наложены траектории аксональных проекций и дендритных полей дисковидных нейронов.
Б. Терминальные зоны проекций от верхней оливы. Возбуждающие проекции от контралатеральной латеральной верхней оливы (ЛВО) доминируют в ростральных областях (закрашенные треугольники).
Возбуждающие проекции от ипсилатерального МВО в основном обнаруживаются в каудальных областях (незакрашенные кружки).
Ингибирующие входящие проекции от ипсилатерального ЛВО также обозначены (синие квадраты). На вкладке показано относительное положение нижнего бугорка внутри центральных проводящих путей слухового анализатора.
Физиологические свойства нейронов центрального ядра нижнего бугорка. Ответы были зарегистрированы у децеребрированных кошек.
Карты частотных ответов представлены на графике зависимости частоты ответных импульсов от частоты и уровня громкости чистого тона.
Условные обозначения графиков представлены на рисунке ниже.
б) Кодирование информации. В исследованиях кодирования информации в нижних бугорков четверохолмия (НБЧ) было определено представление простых элементов звука (амплитуда, частота и время), а также выявлены более сложные структуры, поддерживающие связь, локализацию и группировку восприятия. Направлением этого исследования является изучение ответов, являющихся более избирательными, чем ответы в нижележащих центрах обработки данных. Ответы одиночных нейронов могут быть точно настроены не только на частоты, но и на уровень звукового давления и временные свойства, такие как амплитудная модуляция (AM) или частотная модуляция (ЧМ). Следовательно, в отличие от кодов популяции нейронов слуховой части ствола мозга, ответы отдельных нейронов могут означать критическую биологическую информацию, которая заключается в спектральной форме или динамических паттернах/моделях естественных звуков.
Временное кодирование основано на способности слуховых нейронов устанавливать скорость разряда по варьирующим по времени акустическим свойствам, таким как монауральные колебания AM и ЧМ, а также бинауральные временные различия. Сигналы модуляции являются важными информативными элементами звуковых каналов и основной точкой отсчета для отделения звуковых сигналов от фонового шума. Межушная разница во времени, как описано выше, лежит в основе азимутальной локализации звука.
Амплитудная и частотная модуляции кодируются разницей во времени между потенциалами действия в слуховом нерве и стволе мозга. При тонах низкой частоты нейроны возбуждаются в определенной фазе кривой стимула. У большинства млекопитающих, верхняя граница фазовой синхронизации достигается примерно на частоте 5 кГц. Тем не менее, этот механизм продолжает действовать при значительно более высоких частотах, поскольку скорости разряда синхронизируются с огибающими колебаниями сложных звуков.
Помимо верхних граничных частот фазовой синхронизации и границ последующих ответов, временное кодирование сдерживается вероятностным характером синаптических событий. Каждый синапс добавляет ошибку в нейронное представление времени из-за неопределенности при выпуске нейромедиатора из пресинаптического нейрона и последующей генерации потенциалов действия в постсинаптическом нейроне. Специализированные временные пути, которые проецируются на оливарный комплекс минимизируют эти «колебания» путем интеграции входов многочисленных волокон слухового нерва и надежной передачи этой информации через мощные концевые синапсы.
Дальнейшей синаптической деградации временной информации можно избежать на высоких уровнях слуховой обработки путем преобразования временного кода в код скорости. Нейроны в НБЧ точно настроены на специфическую скорость модуляции, как нейроны ствола мозга настроены на частоту. Следовательно, нейрон действует как «метка» сигнализируя частоту модуляции с ее относительной скоростью импульсации, а не временную структуру своих ответов.
Восходящие проекции латеральной верхней оливы (ЛВО) и медиальной верхней оливы (МВО наделяют нейроны в центральном ядре бинауральной временной настройкой. Как и в стволе головного мозга, большинство этих нейронов показывают ответы пикового и желобообразного типов, что в соответствии с моделью Джеффресса, может быть объяснено бинауральными возбуждающими и тормозными взаимодействиями. Некоторое количество нейронов, однако, показывает чувствительность РПС промежуточного типа. Нейроны могут лучше о твечать на промежуточные различия межушных фаз или отображать комплексные фазочастотные связи. Этот временной признак не совместим с моделью Джеффресса и, вероятно, отражает конвергенцию оливарных входов с различной настройкой РПС. Вполне вероятно, что даже нейроны НБЧ с менее сложными ответами пикового и желобообразного типа объединяют множественные входы из верхней оливы, так как пропускная способность/диапазон частот настройки РПС для обоих типов нейронов значительно точнее, чем в КВО.
Спектральное кодирование в нижних бугорках четверохолмия (НБЧ) является важным для монауральной локализации звуков и речевой коммуникации. Рецепторные поля нейронов НБЧ могут охватывать несколько октав, создавая области, где спектральная энергия вызывает либо возбуждающие, либо ингибирующие эффекты. Звуки с энергией на возбуждающих частотах вызывают сильные ответы; в то время как звуки с энергией на ингибирующих частотах вызывают слабые ответы. Ответ на звуки сложной спектральной формы, такие как широкополосный шум, который фильтруется за счет функции проведения наружного уха, определяется балансом этого возбуждения и торможения.
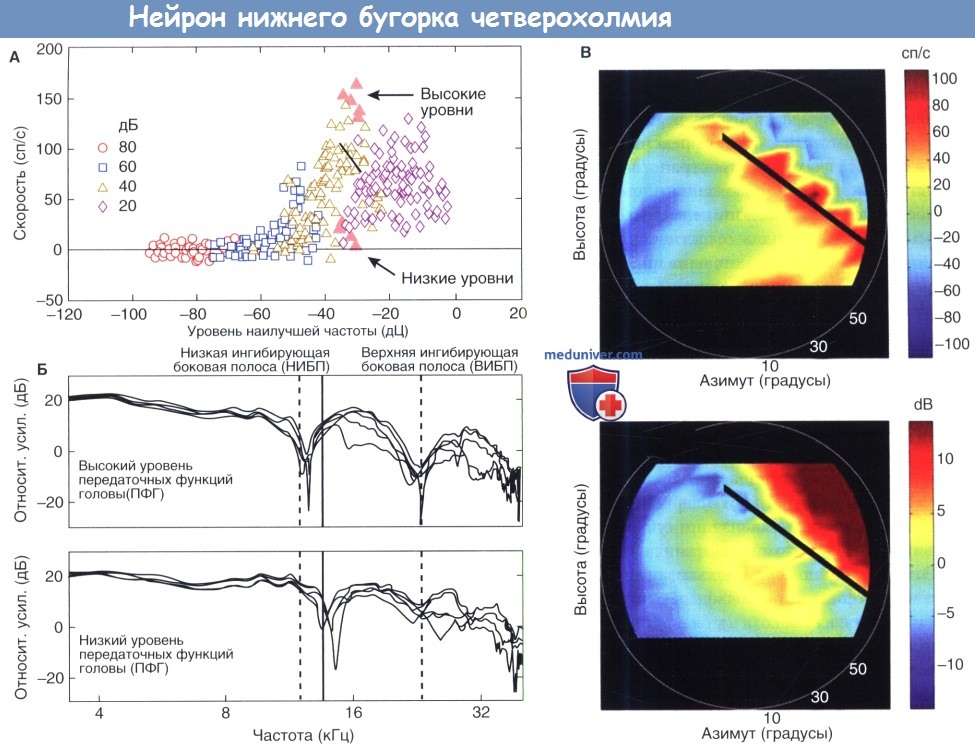
Функциональные результаты сравнения с эталоном приведены на рисунке ниже, показывая, как пространственная настройка нейрона НБЧ коррелирует с частотным диапазоном ВНЕ-НЧ его торможения. На рисунке скорость разрядов нейрона показана для ПФСА-фильтрованных шумовых выбросов. Бинауральные стимулы были представлены через наушники для моделирования акустических эффектов на оба уха в 99 различных местоположениях фронтального звукового поля. Ответы на эти «виртуальные» местоположения звука показаны на четырех уровнях представления. Каждый ответ представлен в соответствии со спектральной энергией в контралатеральном (доминирующем) ухе на НЧ нейрона (12 кГц). Отличительной особенностью полученной функции «скорость-уровень» является крайняя изменчивость в ответах на ПФСА с аналогичной энергией на НЧ. Эти различия наблюдаются, поскольку нейрон является чувствительным к спектральным характеристикам, которые существуют на частотах далеко за пределами НЧ.
ПФСА формы на рисунке ниже сгруппированы по способности вызывать высокую или низкую скорость разряда от нейрона IC, что обозначено закрашенными символами/треугольниками на рисунке. Хотя функции проведения имеют сходную энергию на ВЧ, есть последовательные различия в местоположении частот их выступающих спектральных зубцов. Высокоскоростные ПФСА имеют зубцы, которые находятся в пределах нижних и верхних ингибирующих боковых полос (вертикальные пунктирные линии). Следовательно, они вызывают сравнительно слабые тормозные реакции. Низкоскоростные ПФСА демонстрируют противоположную полярность. Их спектральные зубцы соответствуют центральной области возбуждения, соседней с ВЧ нейронов (вертикальная сплошная линия).
Чувствительность нейрона к формам ПФСА переведена на пространственную регулировку по частоте на рисунке ниже. Верхний график отображает скорости разряда на рисунке в пространственных координатах, которые указывают местоположение виртуального звукового поля стимулов ПФСА. Сильнейшие ответы нейрона (красный) следуют контуру от высокой ипсилатеральной до низкой контралатеральной высоты/уровня (наложенная линия). Нижний график отображает относительный прирост стимулов ПФСА в той же системе координат. Этот прирост избирательно относится к спектральной энергии, находящейся в пределах мощной нижней ингибирующей боковой полосы нейрона (НИБП). Близкое совпадение между местами с малым приростом (синие области на нижнем рисунке) и местами с высокой скоростью (красные области на верхнем рисунке) предполагает, что ингибирование вне-НЧ является основным фактором, определяющим пространственную регулировку нейрона.
Пространственная регулировка/настройка по частоте ответа одного нейрона в нижнем бугорке четверохолмия децеребрированной кошки:
А. Скорость разрядов была вызвана всплеском шумов, что было сформировано передаточной функцией головы кошки (ПФГ, см. дорсальное кохлеарное ядро).
Ответы показаны для 99 местоположений/точек во фронтальном звуковом поле на четырех уровнях представления. Каждая точка данных отображается по уровню спектра на наилучшей частоте нейрона (НЧ).
Закрашенные символы/ треугольники соотвествуют ПФГ с аналогичной на-НЧ энергией, вызванной разницей скорости разряда.
Б. Спектральные формы высокои низкоскоростных ПФГ. Вертикальные линии отмечают относительный прирост функций на-НЧ, на более низкой ингибирующей боковой полосе (НИБП) и верхней ингибирующей боковой полосе (ВИБП).
В. Роль ингибирующей боковой полосы пространственной регулировке/ настройке нейрона. На верхнем рисунке, скорости разряда в А представлены на пространственных координатах.
На нижнем рисунке, прирост ПФГ на НИБП представлены на пространственных координатах. Сильнейшие ответы (красный) наблюдаются, когда функции передачи показывают слабый прирост на ингибирующих частотах.
в) Исследования реакции/абляции. Вклад нижних бугорков четверохолмия (НБЧ) в реакции на локализацию звука был изучен посредством оценки последствий плановых хирургических вмешательств. Значительная часть этой работы включает в себя селективное повреждение одного полушария мозга, что нарушает ряд сложных слуховых функций, полностью не устраняя основной слух. Когда эта процедура выполняется на экспериментальных животных, выявляется поразительное противоречие между поражениями нижних комиссуральных путей и повреждением нижних бугорков четверохолмия (НБЧ). Односторонние поражения на нижних ядрах ствола мозга оказывают негативное влияние на направленный слух, независимо от расположения источника звука. Всеобъемлющий характер дефицита предполагает, что основы локализации прерваны до того, как полностью сформировано нейронное представление информации о направлении. Повреждение любой стороны мозга имеет глобальные последствия, поскольку сроки и величина бинауральных ответов должны объединиться для завершения первого этапа обработки данных о направлении. Напротив, при локализации односторонних поражении в НБЧ, ошибки локализации ограничены контралатеральной половиной слухового поля. Это пространственное ограничение подразумевает второй этап «распределительного» процесса, который смещает представительство более высокого порядка слухового пространства в направлении контралатерального местоположения источников. Как отмечалось ранее, акустический перекрест является предполагаемым механизмом для селективного латерализованого распределения восходящих оливарных входов к центральному ядру. Поэтому повреждение латеральной петли вызывает аналогичные дефициты.
Исследования реакции-абляции в значительной степени полагались на испытания в свободном поле, чтобы охарактеризовать точность данных о направленном слухе. В классической модели двухальтернативного принудительного выбора один из двух ящиков не был заперт, чтобы обеспечивать доступ животных к пищевой награде. Левый, в отличие от правого незапертого ящика, сигнализирует предъявлением звукового раздражителя от подключенного динамика. Пределы возможностей локализации субъекта измеряются изменением угла между комбинациями двух коробок/динамиков. Это подлинная задача локализации, требующая от субъекта идентификации и подхода к источнику звука для пищевого подкрепления.
Обзор классической литературы приводит к впечатлению, что разрушение комиссур НБЧ, либо коры головного мозга не вызывает радикального изменения реакции локализации. Повреждения трапециевидного тела могут вызвать сильный дефицит, но эти эффекты наблюдаются только у некоторых субъектов и они, как правило, преходящи.
Последующие исследования дали возможность проводить тестирование с ящиком с искажеными слуховыми стратегиями, которые не требуют пространственного слуха. Когда два источника звука находятся в левой и правой половине слухового поля испытательной арены, субъект с нормальным пространственным слухом может идентифицировать активный динамик, опираясь на бинауральные сигналы, чтобы отличить левый от правого. Когда пространственная обработка искажается в одной половине слухового поля (например, после поражения левого НБЧ нарушается локализация в правой половине слухового поля), субъект может продолжать определять источник пищи, решая, остался ли ящик с пищей слева или не слева. Когда односторонний дефицит сопровождается потерей слуха, субъект может ответить громко, а не тихо.
Критерии сопряженной изменчивости принятия решения могут быть взяты под контроль стимула за счет увеличения количества источников звука на испытательной арене, которая требует от субъекта выбрать активный динамик из альтернативных местоположений в обеих половинах слухового поля. Это изменение процедуры не влияет на эффекты от поражения трапециевидного тела, но поведенческие последствия поражения вышележащих структур становятся все более очевидными. Без помощи непространственных слуховых стратегий, полное и постоянное нарушение локализации звука наблюдается в контралатеральном звуковом поле.
г) Аудиологические последствия. Признаком анализа звука в НБЧ является четкий баланс возбуждения и подавления. Когда эти процессы функционируют нормально, нейроны настраиваются на многие акустические параметры, что придает смысл звуку. К сожалению, эти механизмы, которые формируют звуковую активность слухового центра среднего мозга, могут выходить из-под контроля у людей, которые испытывают потерю слуха, акустическую травму или старение. Когда возбуждающие импульсы от ствола мозга сокращаются по количеству или силе, тогда НБЧ может ничего не регистрировать. И наоборот, потеря тонического подавления может редуцировать ненормально реагирующие нейроны. Эту гиперактивность рассматривали как причинный фактор аудиогенных слуховых приступов, гиперакузии, усиления громкости, шума.
Иммуноцитохимическое исследование показало значительную потерю подавляющего нейротрансмиттера ГАМК в центральном ядре взрослых крыс. На 1/3 меньше ГАМК негативных нейронов наблюдается в вентролатеральной части ядра контрольной группы. Нейрохимические анализы подтверждают, что вызванное калием высвобождение ГАМК в этих тканях значительно снижено, в то время как высвобождение возбуждающих нейротрансмиттеров глютаматом является повышенным. Эти данные показывают, что нарушенная трансмиссия ГАМК в НБЧ может способствовать неврологическому пресбиакузису. Эффективность ГАМК понижается и при экспериментальных действиях, которые способны вызывать ушной шум.
НБЧ было предложено как момент аудиогенных приступов судорог у животных моделей. На неврологическом уровне индукция эпилептической активности наблюдается в виде ряда длительных следовых реакций, которые следуют за звуковым стимулом. Оказывается, что невосприимчивость к эндогенным ГАМК ингибиторам заложена в ядре этих патологических моделей реакции, когда ГАМК поступает в НБЧ нейрон посредством ионофореза, количество нейротрансмиттеров, которые требуются для подавления активности у склонных к эпилепсии крыс значительно выше, чем у здоровых крыс. О снижении ГАМК-опосредованного подавления также свидетельствует пониженная эффективность агонистов, таких как бензодиазепин. Эпилептическая активность может индуцироваться в норме в нейронах путем ионофореза бикукулина, потенциального антагониста ГАМК.
Эта же стратегия центрального ингибирования может привести к возникновению хронического ушного шума. Помимо продуцирования более сильного, более длительного ответа на слуховой стимул, пониженная регуляция ГАМК может повысить спонтанную активность в отсутствии звука. Так как повышенная активность имитирует вызванные звуком ответы звуковых нейронов, выдвинута гипотеза, что высшие мозговые центры интерпретируют эту активность как звук даже при отсутствии физического стимула. Нижний бугорок четверохолмия (НБЧ) может играть выдающуюся роль в этом явлении по причине сложившейся физиологической уязвимости его ингибированной структуры. Также на дорсальном кохлеарном ядре (ДКЯ), другом предполагаемом источнике шума в ушах, нижний бугорок четверохолмия (НБЧ) получает противоречивые стимулы как от слуховых, так и неслуховых путей. Одна неслуховая функция, то есть инициация условной ответной реакции, может быть продемонстрирована при помощи электрической или химической стимуляции слухового центра среднего мозга. По мере сокращения центрального ингибирования эти подсистемы могут усиливать воспринимаемую тяжесть ушного шума, часто не соответствующую объективным параметрам громкости.
Предполагаемая роль ГАМК при шуме в ушах создает возможность искать способы фармакологических вмешательств. Так как потенциально токсические вещества не могут быть напрямую тестированы на людях, то исследователи должны полагаться на животные модели с экспериментально вызванным ушным шумом. Этот подход ставит нас перед головоломкой, как охарактеризовать субъективное восприятие фантомного звука у животного? Хотя существует несколько поведенческих проверочных процедур, основной стратегией является натаскивание на начало интервала тишины, которая создается при помощи отключения продолжающегося фонового звука. После создания шума под влиянием громкого звука или большой дозы салицилата животное больше не может выполнять задание, так как оно слышит фантомный звук вместо тишины. Успех лекарственной терапии показан посредством восстановления нормальной поведенческой модели. Эти парадигмы в настоящее время указывают на многообещающие результаты применения препаратов, модулирующих трансмиссию ГАМК.