Вестибулярная система собирает информацию о положении и движении головы. Вместе с визуальными и проприоцептивными сигналами мозг использует эту информацию, чтобы координировать глаза, голову и тело во время движения и обеспечивать сознательное восприятие ориентации и движения. Нарушения в этом интеграционном процессе может привести к головокружению, которое является девятой по частоте причиной посещения врачей первичного звена и наиболее распространенной среди пациентов старше 75 лет.
Многих из этих пациентов направляют к отиатру, чтобы определить связь жалоб с вестибулярной патологией и разработать план лечения. Знания анатомии и физиологии вестибулярной системы дает рациональную основу для понимания причин, диагноза и лечения многих видов головокружения.
Разделение вестибулярной системы на периферическую и центральную части помогает уточнить местоположение патологии. Периферический отдел определяет ускорение головы. Все ускорения можно разделить на шесть компонентов: вращение вокруг оси, в каждом из трех измерений и линейное движение по каждой из этих осей.
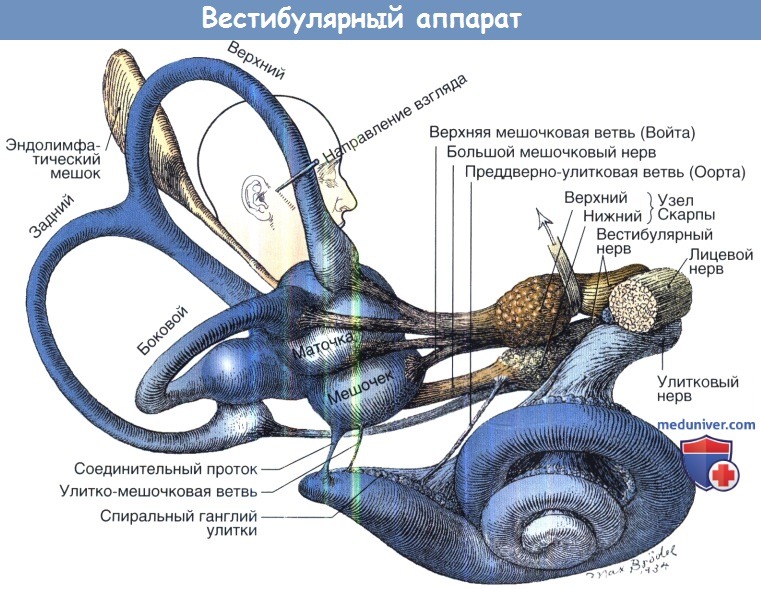
Периферический отдел состоит из горизонтального, верхнего (известного как передний) и заднего полукружных каналов, измеряющих вращение, и utricle (маточки) и saccule (мешочка), измеряющих линейное ускорение. Каждый полукружный канал имеет синергичный канал в противоположной височной кости, расположенный почти параллельно. Горизонтальные каналы действуют как пара, в то время как каждый верхний канал соединен с задним каналом на противоположной стороне.
Линейные силы могут явиться результатом передвижения головы из стороны в сторону, вверх и вниз при изменении ориентации головы относительно центра силы тяжести. Центральная вестибулярная система состоит из бокового, среднего, верхнего и нижнего вестибулярных ядер и их путей к таламусу, коре, мозжечку, спинному мозгу и экстраокулярному двигательному ядру.
Каждый перепончатый полукружный канал заполнен эндолимфой, внеклеточной жидкостью, богатой калием, омываемый перелимфой, которая по составу похожа на цереброспинальную жидкость. Концы каналов открываются в преддверие. Место рядом с концом каждого мембранного канала является расширением, называющимся ампула, которая содержит ампулярный гребешок и куполу.
Структуры лабиринта: мешочек, маточка, передний или верхний полукружные каналы,
задний полукружный канал и горизонтальный или боковой полукружный канал.
Верхний вестибулярный нерв иннервирует горизонтальный, передний полукружный канал и мешочек. Нижний вестибулярный нерв иннервирует задний полукруглый канал и мешочек.
Клетки для вестибулярных нервов находятся в вестибулярном ганглии. Рисунок из архивов Brodel.
Ампулярный гребешок представляет собой имеющую форму седла желеобразную структуру вдоль одной из стен мембранного канала, которая содержит волосковые клетки вестибулярной системы. Купула действует как мембранозная диафрагма, простирающаяся от кристы к противоположным стенкам канала. В связи с привязкой мембранных лабиринтов к черепу гребешок и купол ускоряются вместе с головой при вращении, но из-за инерции эндолимфы происходит отставание.
Эндолимфа накапливается с более высоким давлением на одной стороне куполы, чем на другой. После прекращения ускорения головы купол и гребешок постепенно возвращаются в положение покоя.
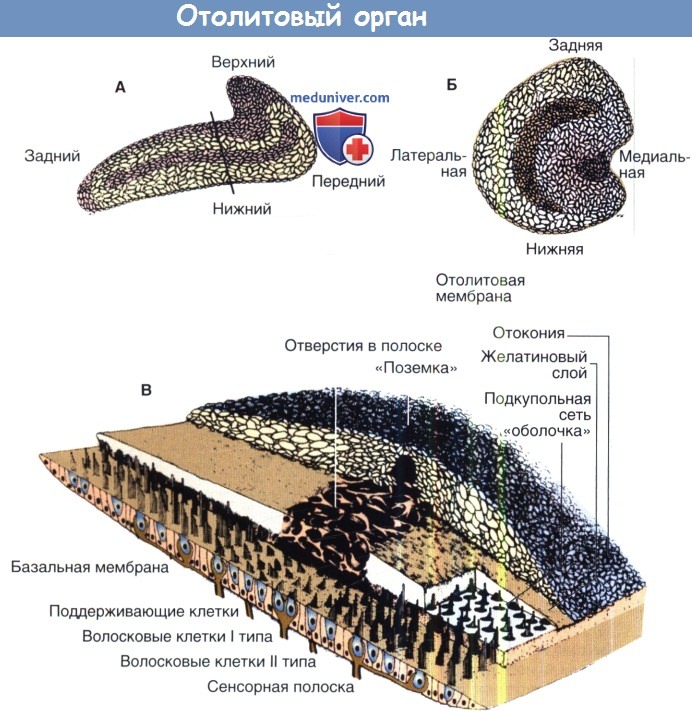
Маточка и мешочек, составляющие отолитовый аппарат внутреннего уха, также содержат волосковые клетки, которые изгибаются в ответ на ускорение головы. Вместо гребешка чувствительный эпителий этих органов покрыт плоскими почковидными слоями, которые называются пятнами. Одно пятно расположено горизонтально в верхней части мешочка, остальные по стенкам. Каждая макула является желатиновой матрицей, с захождением волосковых клеток и покрытием крошечными гранулами углекислого кальция — отолитами.
Пятна отолитового органа чувствительны к линейному ускорению из-за присутствия отоконий на их поверхности. Они увеличивают плотность пятен так, что при линейном движении головы тяжелые пятна отстают, сгибая встроенные в них волосковые клетки. Маточка в первую очередь чувствует боковой наклон и перемещение головы, в то время как мешочек измеряет движение вперед-назад и наклон перевода, а также движения в соответствии с действием силы тяжести.
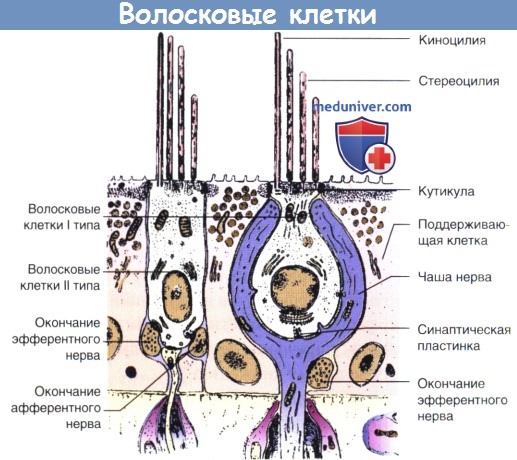
Волосковые клетки, расположенные в сенсорном эпителии полукружных каналов, и отолитовые органы несут ответственность за кодирование ускорения головы в модуляции разряда скорости вестибулярных афферентных волокон. Каждая волосковая клетка содержит комплект из 50-100 стереоцилий, и одну длинную киноцилию на краю каждого пучка (рис. 6-5). Расположение этой киноцилии по отношению к стереоцилии дает каждой волосковой клетке внутреннюю полярность.
В горизонтальных каналах киноцилия каждой волосковой клетки находится на стороне цилиарного расслоения перед мешочком, в то время как этот механизм будет обращаться вспять в верхнем и заднем каналах. Волосковые клетки каждого отолитового органа расположены в двух полосах, расположенных вдоль центральной полосы, которые называются striola (стриолами). Киноцилии из волосковых пучков в мешочке ориентированы на стриолу, а она на поверхность мешочка.
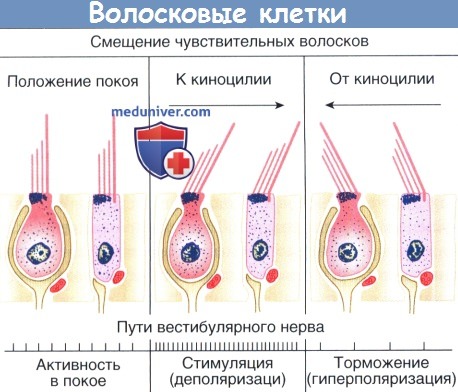
Смещение цилиарного расслоения к своей киноцилии открывает калиевые каналы вдоль ресничек и деполяризует волосковые клетки от потенциала покоя мембраны до -50-(-70) мВ. Чувствительность волосковых клеток может приблизиться к 20 мВ деполяризации в микрометре перемещения. Эта поляризация приводит к притоку кальция в базальном конце волосковых клеток и увеличению потока нейромедиаторов в синапс, в то время как перемещение в обратном направлении гиперполяризует клетки и уменьшает освобождение нейромедиатора.
Ориентацию киноцилий из полукружных каналов определяет возбужденное направление каналов.
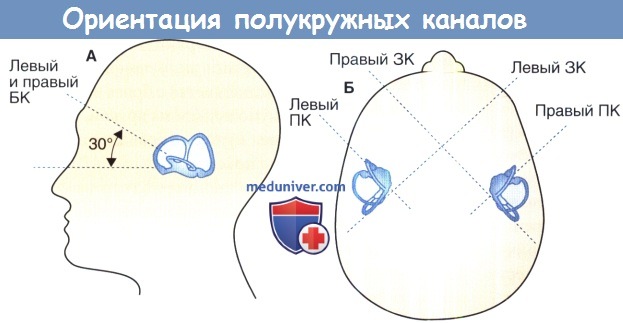
Ориентация полукружных каналов:
А. Горизонтальный (боковой) полукружный канал.
Б. передний (верхний) и задние каналы.
БК—боковой канал, ПК—передний канал, ЗК — задний канал.
Относительные размеры каналов преувеличены для наглядности.
Каждый горизонтальный канал максимально возбуждается при вращении в сторону канала и подавляет вращение в обратном направлении. Это приводит к возбужденной фазе медленного движения в сторону противоположного канала и сброс саккады к каналу. Верхний канал возбуждается вращением вниз и в стороны в плоскости канала. Это приводит к вертикально-ротаторному нистагму, с медленной фазой и вертикальной составляющей вверх, и позволяет сбросить саккады вниз. Задний канал возбуждается от вращения вверх и в стороны в плоскости канала, так что медленная фаза стремится вниз.
Афферентные нервные волокна в верхнем вестибулярном нерве простираются от сенсорных структур в верхнем и горизонтальном каналах и мешочке к вестибулярным ядрам в стволе головного мозга. Нижний вестибулярный нерв идет от заднего полукружного канала и мешочка. У людей каждый вестибулярный нерв состоит примерно из 25000 нейронов. Эти нейроны являются биполярными, т.к. импульс передается с клеточных тел, расположенных в вестибулярном нерве вблизи ствола мозга, в вестибулярный ганглий. В каждом лабиринте помимо афферентных от вестибулярных ядер к волосковым ядрам идут примерно 400-600 эфферентных нервов.
Точная роль этих эфферентов в модуляции физиологии вестибулярного рефлекса неизвестна, но может быть связана с регулировкой чувствительности вестибулярного аппарата к предстоящему волевому движению.
Почти все вестибулярные афференты нерва имеют спонтанные или свободные разряды, с некоторыми волокнами возбуждения до 100 колебаний/сек. Этот покой в разряде позволяет каждому из афферентов реагировать как на возбуждающие, так и на тормозные раздражители. Разрядом регулярности измеряется расстояние между потенциалами действия разряда афферентов, что предоставило полезный маркер для многих важных физиологических процессов. Три группы вестибулярных афферентных нервов были выявлены у млекопитающих на основе их ответов на движение и их профилей иннервации в рамках сенсорного эпителия лабиринта.
Только афферентные окончания синапсов имеют включения на волосковых клетках типа II в периферийной зоне кристы. Эти афференты регулярно разгружаются и имеют низкий уровень чувствительности к вращению. Диморфные афференты имеют чашевидные окончания, заканчивающихся в I типе волосковых клеток, и окончаний синапсов, заканчивающиеся в волосковых клетках типа II. Их физиологические свойства меняются в зависимости от расположения на гребешке.
Такие диморфные афференты заканчивающиеся в периферической зоне, регулярно разряжаются, а заканчивающиеся вблизи центральной зоны освобождаются неравномерно при более высокой чувствительности к вращению. Существует также группа афферентных волокон доходящих только до чашечки окончаний на волосковых клетках I типа в центральной зоне. Заканчивающиеся только у чашечки афферентные волокна имеют неравномерный разряд и обладают низкой чувствительностью к вращению при низкой частоте стимула и высокой при высокой частоте. Функция различных типов волокон может быть связана с каждым типом движения головы.
В выполненной более века назад работе Эвальд определил два основополагающих принципа, регулирующих отношения между лабиринтами, рецепторами и передаваемыми ими вестибулярными рефлексами. При экспериментах на голубях он установил, что фенестрация полукружных каналов с последующей механической стимуляцией мембранного канала приводит к движению глаз и головы, которые были в плоскости этого канала.
Он также отметил, что возбуждающие стимулы привели к большей амплитуде, чем ответы тормозных раздражителей. Это, по крайней мере частично, объяснимо разрядом вестибулярных афферентных нервов и центральных вестибулярных нейронов в покое. Эти нейроны могут быть возбуждены до по меньшей мере 350 колебаний/сек, но возможно и торможение до 0 колебаний/сек. Таким образом, диапазон возбуждения в три-четыре раза выше, чем для торможения. Многие центральные вестибулярные нейроны имеют разряд покоя, предполагая, что их ответ может также способствовать асимметрии в ответной реакции.
Движения глаз и головы в плоскости пострадавшего канала, называются первым законом Эвальда. Второй закон Эвальда гласит: «возбуждающих ответов больше, чем тормозящих». Эти отношения обеспечивают основу для понимания многих симптомов и признаков, которые происходят после травмы лабиринта.
Структура ампулярной кристы и куполы:
А. Эскизное изображение ампулярного гребешка. Б. поперечное сечение ампулярной кристы обезьяны.
Гистологический препарат: место прикрепления купола от верхушки гребешка до противоположной стороны перепончатой ампулы.
Стрелками указано пространство под куполом.
Структура отолитового органа.
А. мешочек. Б. маточка.
В. Состав мембраны отокония у мешочка в разрезе, что показано на уровне А.
Типы волосковых клеток (типы I и II) находятся в лабиринте млекопитающих.
Распространение путей вестибулярного нервного волокна в зависимости от смещения ресничек.
Вестибулярные афферентные нервы возбуждаются чаще, когда волоски пучками смещаются в сторону киноцилии,
они стреляют медленнее, когда смещаются в противоположном направлении.
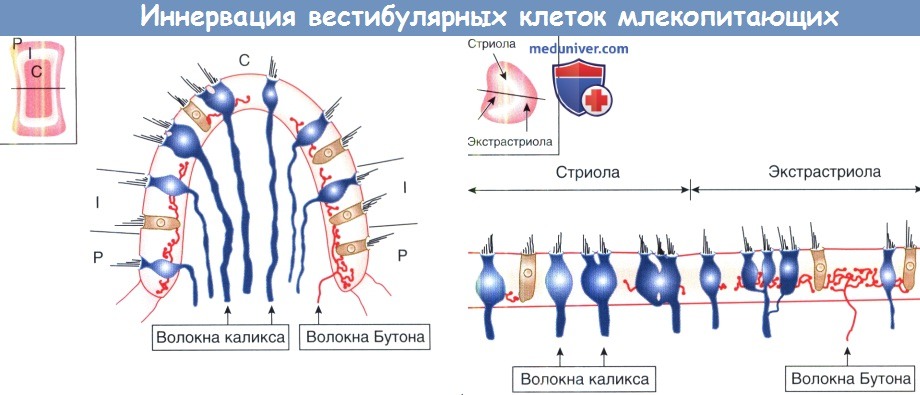
Афферентные модели иннервации вестибулярных конечных органов млекопитающих.
Верхний рисунок: нейроэпителий крист делится на центральную (С), промежуточную (I) и периферическую (Р) зоны, как показано на плане на вставке и в поперечном сечении на главной панели.
Диморфные волокна находятся повсюду. Нижний рисунок: макулы делятся на striola (стриолы) и латеральные и медиальные extrastriola (экстрастриолы).
Чашечка и волокна находятся в striola, волокна Бутона в extrastriola.