а) Бинокулярная декорреляция любой этиологии запускает каскад латентного нистагма. Клинические исследования детей и взрослых с латентным нистагмом инспирировали серию поведенческих, физиологических и нейроанатомических исследований на приматах с латентным нистагмом, сопровождающимся инфантильным косоглазием, возникшим в силу естественных причин, или вызванным в ходе эксперимента. Обычно полученные результаты свидетельствовали о том, что частота и тяжесть латентного нистагма коррелируют с возрастом дебюта и длительностью существования бинокулярной декорреляции в младенческом возрасте.
Наиболее частая клиническая причина бинокулярной декорреляции — косоглазие, которое у младенцев человека в подавляющем большинстве случаев сходящееся (эзотропичное, конвергентное). Рано дебютирующая эзотропия встречается чаще эзотропии, их соотношение составляет 9:1. Эзотропия также является наиболее часто встречающейся формой естественным образом развивающегося косоглазия у приматов.
Однако длительная депривация бинокулярных стимулов любой этиологии в раннем младенческом возрасте (монокулярная врожденная катаракта, односторонняя дальнозоркость, близорукость или астигматизм высокой степени, кровоизлияние в стекловидное тело одного глаза у новорожденного, помутнение роговицы на одном глазу, плотные двусторонние катаракты, моделирование этого состояния у обезьян) могут вызывать бинокулярную декорреляцию; причиной может быть, в том числе, и монокулярная депривация (односторонняя амблиопия) или тяжелая бинокулярная депривация (двусторонняя амблиопия), а также тарзорафия (тонкие полупрозрачные веки обезьян имитируют врожденную катаракту, создавая диффузное освещение сетчатки, но препятствуют предметному зрению).
Утрата предметного зрения не является обязательным условием; у большинства младенцев людей и младенцев обезьян с косоглазием изначально отсутствует амблиопия и наблюдается альтернирующая фиксация. Необходимым и достаточным фактором является бинокулярная декорреляция, а не низкая острота зрения.
Длительность существования декорреляции, превышающая период, эквивалентный трем месяцам развития младенца человека, в 100% случаев приводит к латентному нистагму. Дефект входящих путей на первой неделе жизни вызывает развитие латентного нистагма, но также латентный нистагм развивается при возникновении нарушений и в момент возникновения нормальной фузии и стереопсиса (эквивалент 2-4-месячному возрасту младенцев человека). Тяжесть развившегося латентного нистагма коррелирует со степенью нарушения бинокулярных связей между глазными доминантными колонками разных глаз зоны V1 зрительной коры и выраженностью интерокулярной супрессии.
Из зоны VI идут связи в экстрастриарные зоны (зоны МТ/MST — медиальной височной/медиальной верхневисочной коры), играющие важную роль в фиксации взгляда и движениях глаз, в том числе плавных следящих движениях, оптокинетическом нистагме и коротколатентных реакциях слежения глаз.
б) Нарушения в зоне V1 вызывают дисфункцию в медиальной височной/медиальной верхневисочной коре. Зрительные зоны V1, V2 (престриарная кора), МТ (medial temporal — медиальная височная) и MST (medial superior temporal — височная верхнемедиальная) коры головного мозга являются основными компонентами пути содружественных движений глаз. У здоровых приматов каждая из этих зон содержит пространственно селективные бинокулярные нейроны. Медиальная височная кора каждого полушария регулирует направление взора ипсиверсивного глаза. От височной верхнемедиальной коры идут нисходящие связи к глазодвигательным ядрам ствола, управляющим движениями глаз: ядру зрительного тракта (NOT), медиальному вестибулярному ядру и взаимосвязанным ядрам отводящего и глазодвигательного нервов. Возможно, у приматов роль субкортикальных входящих путей к ядру зрительного тракта невелика. Основной путь — церебральный, от височной верхнемедиальной коры к стволу головного мозга.
Доминирующая роль кортикального пути и минимальное значение субкортикального пути подтверждается исследованиями детей. С помощью лучевых методов исследования зрительной коры в комбинации с окулографией удалось установить, что у младенцев с церебральной слепотой отсутствуют следящие движения или оптокинетический нистагм, вызываемые зрительной стимуляцией.
Предполагалось, что одним из механизмов возникновения асимметрии взора может быть преобладание «носовых» нейронов в зоне V1 зрительной коры и их влияние на медиальную височную зону при незрелости коры и при косоглазии. Однако исследования ответов нейронов зон VI, V2 и медиальной височной коры на движущиеся в разном направлении и бинокулярные стимулы младенцев обезьян, а также обезьян с рано дебютировавшим косоглазием не выявили преобладания нейронов, селективно реагирующих на движения в носовом направлении. У животных с косоглазием отмечается ослабление бинокулярных (возбуждающих) ответов и усиление интерокулярной супрессии. Эти физиологические аномалии имеют нейроанатомический субстрат. В зоне V1 обезьян с косоглазием отмечается недостаточность бинокулярных связей и подавление метаболической активности интерокулярных структур.
в) Бинокулярная декорреляция демаскирует врожденные монокулярные носовые отклонения. Латентный нистагм всегда связан с аномальным бинокулярным развитием в младенческом возрасте. Это важное клиническое наблюдение инспирировало упомянутые выше исследования приматов, в результате которых, в свою очередь, для объяснения патофизиологии латентного нистагма возникла необходимость в исследовании функционально-структурных корреляций. Результаты исследований обезьян позволили детским офтальмологам обосновать возможно более раннее восстановление фузии в младенческом возрасте с целью предотвращения латентного нистагма или уменьшения его тяжести.
Латентный нистагм вызывается дефектом афферентного бинокулярного зрительного пути. Бинокулярный дефект демаскирует асимметрию направления взора, регулируемого церебральными путями взора. При нормальном бинокулярном развитии (фузии) — в первые месяцы жизни — асимметрия направления взора подавляется; аномальное развитие (нарушение развития фузии) усиливает асимметрию. Если в младенческом возрасте фузия не восстанавливается, асимметрия взора сохраняется на протяжении всей жизни.
Зрительная кора каждого полушария головного мозга с рождения тесно связана со структурами, осуществляющими приведение глаза. Эти врожденные связи монокулярны. Чтобы стимулировать фиксацию взгляда при повороте глаза в височную сторону, импульсация должна пройти через бинокулярные связи и не встретить препятствия в виде интерокулярной супрессии. Если не развивается нормальной бинокулярности, система остается преимущественно монокулярной и асимметричной, не способной фиксировать взор в височном направлении или осуществлять устойчивые следящие движения/оптокинетический нистагм в височном направлении. Латентный нистагм — аномальная монокулярная асимметрия, наложившаяся на нормальную ипсиверсивную полушарную асимметрию взора.
г) Гипотетический путь латентного нистагма. На рисунке ниже проиллюстрирован механизм латентного нистагма, показаны пути, регулирующие фиксацию взора у приматов и роль бинокулярных связей. Менее активные вследствие окклюзии и/или интерокулярной супрессии зрительные и моторные нейроны изображены на голубом фоне. В левой половине рисунка показаны пути и структуры зрительно-моторной системы примата с латентным нистагмом.
Импульсация на схеме идет сверху вниз, от монокулярного поля зрения фиксирующего (или зрячего) правого глаза. Носовые и височные части поля зрения у приматов имеют неодинаковую площадь, отклонение глаза к носу способствует работе более обширной височной части поля зрения. Волокна ганглиозных клеток носовой и височной половин сетчатки перекрещиваются в зрительном перекресте, оканчиваются синапсами в наружном коленчатом ядре и проецируются в чередующихся монокулярных зрительных доминантных колонках правого и левого глаза зоны V1. Во время развития ганглиозные клетки носовой части сетчатки становятся более многочисленными и раньше формируют связи, чем ганглиозные клетки височной части сетчатки (не показаны). Пластинки наружного коленчатого ядра, соответствующие носовой части сетчатки — пластинки 1, 3 и 5 (также не показаны) — содержат больше нейронов и развиваются раньше, чем пластинки, связанные с височной половиной сетчатки, а именно пластинки 2, 4 и 6.
В пределах наружного коленчатого тела нейроны остаются монокулярными, бинокулярные интерламинарные соединения отсутствуют.
Монокулярная асимметрия, проявляющаяся более сильной импульсацией от носовой части сетчатки, проецируется на глазные доминантные колонки зоны V1. В зоне V1 каждого полушария глазные доминантные колонки, представляющие носовую часть сетчатки (височную половину поля зрения), занимают немного большую площадь коры (на схеме они изображены более крупными), чем представляющие височную часть сетчатки (носовая половина поля зрения), но каждая глазная доминантная колонка также содержит нейроны, более чувствительные к движению видимого объекта в сторону носа (в данном случае, вправо), чем в височном направлении (влево). На схеме нейроны рецептивных полей зоны V1 и медиальной височной коры для простоты изображены в виде полукругов, обозначающих соответствующие им части поля зрения.
Стрелками отмечена более высокая чувствительность нейронов к движению стимула в ту или другую сторону. Нейроны зрительной зоны (включая нейроны, лежащие за пределами зоны VI в медиальной височной коре) чувствительны к движениям направленным и к носу, и к виску, но лишь нейроны, кодирующие движение в сторону носа с рождения через монокулярные связи связаны с нейронами взора (глазодвигательными) зоны височной верхнемедиальной коры (объединенных в височную верхнемедиальную дорсальную порцию, MSTd — medial superiotemporal dorsal). Височные верхнемедиальные дорсальные нейроны каждого полушария кодируют ипсиверсивные движения глаза, то есть, относительно контралатерального глаза — в носовую сторону (влево — нейроны медиальной верхневисочной коры левого полушария головного мозга и вправо — нейроны медиальной верхневисочной коры правого полушария).
Единственная разница между зрительной корой примата с латентным нистагмом и зрительной корой здорового примата заключается в малом количестве бинокулярных горизонтальных связей (вследствие интерокулярной супрессии). На рисунке недостаточность бинокулярных связей обозначена как отсутствие диагональных связей глазных доминантных колонок левого глаза с глазными доминантными колонками правого глаза в коре примата с латентным нистагмом (левая половина рисунка) и их наличие в коре здорового примата (правая половина рисунка). В коре здоровых приматов для взгляда в височную сторону требуется доступ к височным верхнемедиальным дорсальным нейронам (MSTd) через бинокулярные связи с гомоверсивными нейронами соседних глазных доминантных колонок, представляющих парный глаз (при рассмотрении правого глаза это нейроны глазных доминантных колонок левого глаза). Путь от зоны V1/медиальной височной коры к височной верхнемедиальной дорсальной коре требует эфферентных проекций через валик (splenium) мозолистого тела.
Височные верхнемедиальные дорсальные нейроны (MSTd) отдают нисходящую проекцию в ипсилатеральное ядро зрительного пути (NOT) ствола головного мозга и ипсиверсивные стволовые структуры (медиальное вестибулярное ядро, долсолатеральное ядро моста и глазодвигательные ядра третьего и шестого черепных нервов).
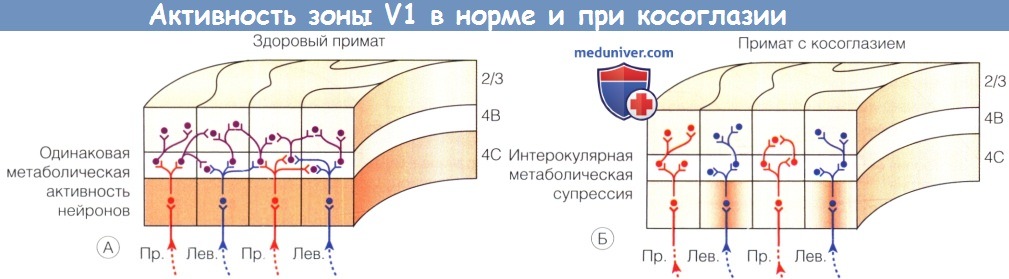
Метаболическая активность соседних глазных доминантных колонок зоны V1 здорового и косоглазого примата.
(А) У здоровых приматов слой 4С при окраске на метаболический энзим цитохром-оксидазу прокрашивается равномерно (отмечено коричневым), что свидетельствует об одинаковой активности колонок правого и левого глаз.
(Б) У приматов с косоглазием в узкой монокулярной зоне в пределах глазных доминантных колонок доминантного глаза (на рисунке—левого) наблюдается нормальная метаболическая активность (отмечена коричневым цветом),
но доминантные глазные колонки подавляемого глаза (на рисунке—правого) и пограничные бинокулярные зоны между доминантными глазными колонками светлые, что указывает на аномально низкую- т.е. подавляемую—активность.
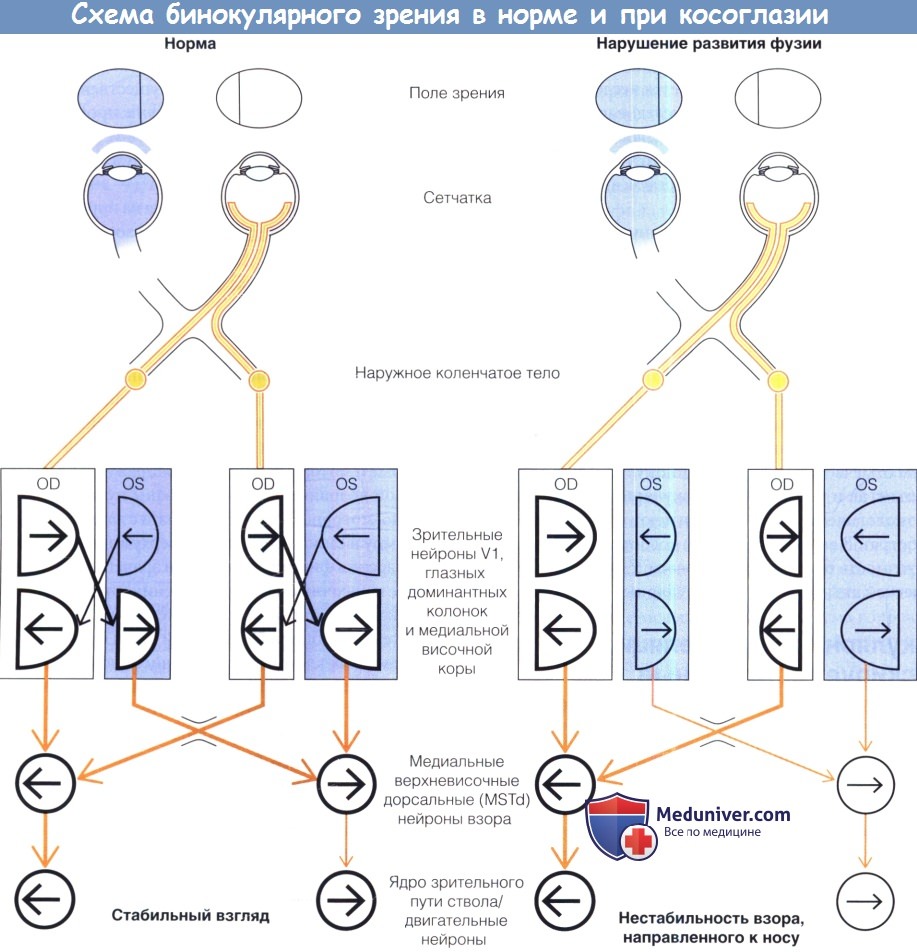
Схема нейронной сети, иллюстрирующая путь зрительного сигнала при слежении и фиксации взгляда у здоровых приматов и при косоглазии.
Недостаток зрелых бинокулярных связей объясняет асимметрию реакций, проявляющуюся асимметрией движений при слежении/оптокинетическом нистагме и латентном фиксационном нистагме.
У всех приматов нейроны зоны слежения каждого полушария кодируют слежение в ипсилатеральном направлении.
Путь сигнала начинается с движущегося стимула в монокулярном поле зрения, который вызывает ответ нейронов зрительной зоны (т.е. зоны V1/медиальной височной коры).
При рождении каждый глаз имеет связь—через врожденные монокулярные связи—с нейронами зоны слежения (например височными верхнемедиальными дорсальными нейронами, MSTd) контралатерального полушария.
Доступ к нейронам слежения ипсилатерапьного полушария требует наличия зрелых бинокулярных связей.
Косоглазие/нестабильность взора, направленного к носу: сверху вниз, начиная от движения объекта в монокулярном поле зрения правого глаза.
Волокна ганглиозных клеток носовой и височной частей сетчатки (глаз) перекрещиваются в хиазме (chi), образуют синапсы в наружном коленчатом ядре (LGN) и отдают проекцию к чередующимся полоскам глазных доминантных колонок зоны V1 (зрительные зоны обозначены прямоугольниками).
В каждой зоне V1 глазные доминантные колонки, представляющие носовую часть сетчатки (височная часть поля зрения) занимают немного большую площадь коры, чем колонки, представляющие височную часть сетчатки (носовая часть поля зрения), но каждая глазная доминантная колонка содержит нейроны, более чувствительные к движениям объекта в носовом направлении, а не в височном (форма полукруга соответствует форме изображаемой части поля зрения, стрелками отмечена более высокая чувствительность нейронов к движению стимула в ту или другую сторону).
Нейроны зрительной зоны (в том числе лежащие запределами зоны V1 в области медиальной височной коры) чувствительны к движению стимула и в носовом, и в височном направлении,
но только нейроны, кодирующие движение стимула в носовом направлении с рождения связаны—через монокулярные связи — с зоной слежения.
Норма/стабильный взор: имеются бинокулярные связи между нейронами с одинаковой ориентацией/более чувствительных к движению стимула в одном направлении глазных доминантных колонок,
представляющих разные глаза (диагональные линии между колонками).
Относительно правого глаза зрительные нейроны, более чувствительные к движениям в носовом направлении, проецируются в зону слежения левого полушария; зрительные нейроны,
более чувствительные к движению стимула в височном направлении, проецируются в зону слежения правого полушария.
Нейроны зрительной зоны, более чувствительные к движению стимула в височном направлении получают доступ к нейронам зоны слежения только через бинокулярные связи.
Call — corpus callosum, через него нейроны зрительной зоны каждого полушария проецируются в контралатеральную зону слежения.
Жирные линии — активные нейроны и нейрональные проекции.