• Транспортные везикулы перемещают белки и другие макромолекулы из одного ограниченного мембраной компартмента в другой, по механизму экзоцитоза и эндоцитоза
• Окаймление из цитоплазматических белковых комплексов помогает образовываться транспортным везикулам и отбирать белки, которые должны быть перенесены
• Белки, предназначенные для транспорта в какой-либо компартмент, отсортировываются от резидентных белков и белков, предназначенных для транспорта в другие компартменты
• Транспортные везикулы используют фиксаторы и SNARE для причаливания и слияния с очередными компартментами на пути транспорта
• Антероградный (прямой) транспорт везикул компенсируется их ретроградным (обратным) транспортом. Транспортируемые везикулы при ретроградном транспорте содержат рециклируемые и сохранившиеся интактными белки
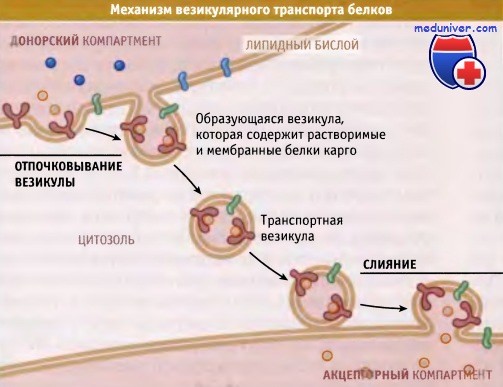
При везикулярном транспорте везикула отпочковывается
от мембраны одного компартмента и сливается с мембраной другого компартмента.
Мы уже отмечали выше, что клетки эукариот содержат множество компартментов, ограниченных мембранами, каждый из которых характеризуется своим составом и функциями. Эти компартменты часто обмениваются компонентами, особенно при эндоцитозе и экзоцитозе. Поскольку обычно белки и липиды свободно диффундируют вдоль липидного бислоя, любое прямое физическое взаимодействие между разнородными мембранными системами создает хаотичность.
Мы выдвинули представление, согласно которому обмен компонентами происходит при участии транспортных везикул, и для поддержания постоянства состава компартмента везикулы должны содержать только отобранные компоненты. В этом разделе мы представим механизм везикулярного транспорта.
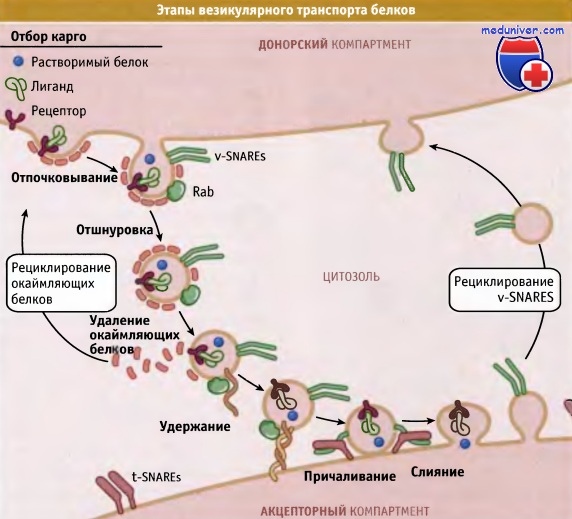
Везикулярный транспорт может быть представлен в виде последовательности нескольких процессов. Эти процессы включают выбор груза (карго), образование везикулы и ее отпочковывание, удаление (окаймления) белкового покрытия, удержание везикулы и ее причаливание (прикрепление), слияние и рециклизацию белков слияния.
Вначале транспортируемые белки должны быть отобраны. На цитоплазматическом конце молекул мембранных белков, например рецепторов, находятся различные сигналы сортировки. Растворимые белки или отбираются соответствующим рецептором, или неизбирательно диффундируют в образующуюся везикулу. Процедуру отбора проходят также белки, используемые на поздних стадиях транспорта. Некоторые резидентные белки, включая мембранные и растворимые, обладают специфическими последовательностями, определяющими их местонахождение в конкретном компартменте. Эти последовательности исключают возможность отбора белков или их заключения в образующуюся транспортную везикулу.
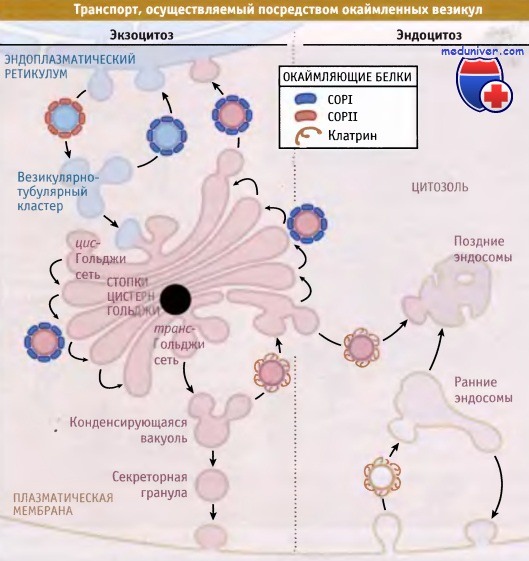
Выбор карго происходит при связывании с сигналами сортировки особых цитоплазматических белков, называемых белки окаймления. Эти белки связываются с сигналами сортировки непосредственно или через адаптерный комплекс, который связывает белок карго с комплексом окаймляющего белка. Последний определяет тип транспортной везикулы. В различных механизмах транспорта участвуют различные белки. Например, белки COPI и СОРИ принимают участие в экзоцитозе, а клатриновые везикулы представляют собой единственный тип везикул, участвующих в эндоцитозе. Состав везикул, которые обеспечивают транспорт между компартментами, отражает лежащий в его основе принцип селективного переноса.
Отличаются друг от друга не только карго белки, но также система их отбора, отпочковывания везикулы, ее причаливания и слияния. Все это определяется донорским и акцепторным компартментами.
Для образования везикулы мембрана органеллы должна образовать выпячивание наподобие почки. В этом процессе принимают участие окаймляющий и адаптерный комплексы, вероятно, связываясь с фосфолипидами, содержащими фосфоинозитол. После образования почки белки «отшнуровки» высвобождают окаймленную белком везикулу, содержащую поглощенный материал. Процесс отшнуровки везикулы требует замыкания мембраны, в результате которого везикула отделяется от донорского компартмента. Механизмы, вызывающие выпячивание мембраны и отшнуровку везикулы, остаются неизвестными.
По сравнению с другими типами везикул, процессы образования и отшнуровки клатриновых везикул исследованы гораздо лучше.
После образования везикулы белки окаймления удаляются (процесс называется «раздеванием») и используются в повторных циклах отпочковывания новых везикул. Освобождение везикулы от окаймляющего белка, по-видимому, необходимо для ее взаимодействия с мембраной реципиентого компартмента.
Присоединение везикулы к реципиентному компартменту происходит в два этапа: первый этап называется удержание, второй — причаливание (прикрепление). Предполагается, что удержание дает возможность везикуле протестировать мембрану, для того чтобы определить, является ли она правильной мишенью. При причаливании мембраны вступают в тесный контакт, обеспечивающий их слияние. В удержании везикулы участвуют небольшие ГТФазы семейства Rab и специальные удерживающие белки, образующие белковые комплексы. Эти комплексы специфичны для каждой пары: транспортная везикула—органелла назначения и таким образом, играют вспомогательную роль в обеспечении адресной доставки материалов.
При удержании белки SNARE, находящиеся на везикуле, образуют комплекс с белками поверхности реципиентной органеллы. Этот комплекс «прикрепляет» везикулу и играет существенную роль в слиянии, которое необходимо для того, чтобы везикула полностью выполнила свою задачу по доставке мембранных компонентов и растворимых молекул к месту назначения. Только при слиянии мембран содержимое двух компартментов может войти в контакт. Многочисленные события слияния мембран, происходящие при экзоцитозе и эндоцитозе, определяются различными комбинациями белков SNARE.
В дальнейшем АТФаза NSF и связывающий белок SNAP разрушают комплексы SNARE, которые образовались при слиянии мембран. Затем из донорской мембраны SNARE поступают в отпочковывающиеся транспортные везикулы и опять возвращаются в донорскую мембрану.
Исследования молекулярных механизмов везикулярного транспорта во многом способствовали выработке новых взглядов по всем аспектам мембранного транспорта. Использование методов реконструкции процессов транспорта или слияния органелл позволило идентифицировать и выделить индивидуальные белки (наприВьщеление мутантов дрожжей (Saccharomyces cerevisiae), дефектных по процессу секреции белка, подтвердило физиологическую значимость этих белков и позволило открыть многие другие белки. В результате проведения исследований по сайт-специфическому мутагенезу и поиску по базам данных были выяснены сигналы сортировки, которые направляют белки в специфические транспортные везикулы.
Процесс везикулярного транспорта состоит из нескольких этапов,
обеспечивающих селективное включение белков карго в транспортные везикулы
и их селективное слияние с мембраной-мишенью.
Три основных типа покрытий, характерных для процесса везикулярного транспорта,
представляют собой COPI, COPII и клатриновое окаймление.