Механизм возврата резидентных белков эндоплазматического ретикулума (ЭПР)
• Присутствующие в большом количестве растворимые белки эндоплазматического ретикулума (ЭПР) содержат последовательности (например, такие как KDEL). За счет соответствующих рецепторов эти последовательности обеспечивают им возможность возврата из поздних компартментов
• Резидентные белки мембран и циклирующие белки возвращаются в ЭПР за счет сигнала из диаминомонокарбоновых кислот, который находится в цитоплазматическом участке молекулы
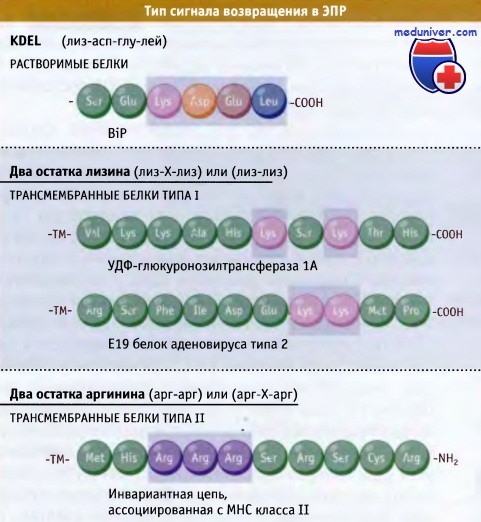
• Сигнал возвращения в ЭПР для трансмембранных белков типа I представлен двумя остатками лизина, а для белков типа II — двумя остатками аргинина
Содержание белков в люмене эндоплазматического ретикулума (ЭПР) составляет более 40 мг/мл. Эти белки представлены вновь синтезированными секреторными белками и большим количеством резидентных белков ЭПР, главным образом шаперонами, способствующими приобретению секреторными белками нативной структуры.
С учетом большого количества этих белков и вероятности того, что, по крайней мере, часть экспорта из ЭПР происходит за счет неизбирательного потока, клетка постоянно рискует «потерять» из ЭПР белки, необходимые для его функционирования. Действительно, такой экспорт происходит, однако он редко приводит к выходу резидентных белков ЭПР во внеклеточную среду. Это связано с существованием эффективного «механизма возврата», благодаря которому белки, вышедшие из компартмента на путь экзоцитоза, возвращаются в ЭПР.
Наиболее полно изучен механизм возврата, связанный с существованием сигнала сортировки, состоящего из четырех аминокислотных остатков. Этот сигнал расположен на С-терминальном участке большинства растворимых резидентных белков люмена ЭПР. В клетках млекопитающих сигнал возвращения представляет собой последовательность Лиз-Асп-Глу-Лей (KDEL), которая впервые была обнаружена в шаперонах. Во всех остальных клетках эукариот обнаружены различные варианты этой сигнальной структуры. У почкующихся дрожжей сигнал возвращения белков в ЭПР представлен последовательностью Гис-Асп-Глу-Лей (HDEL).
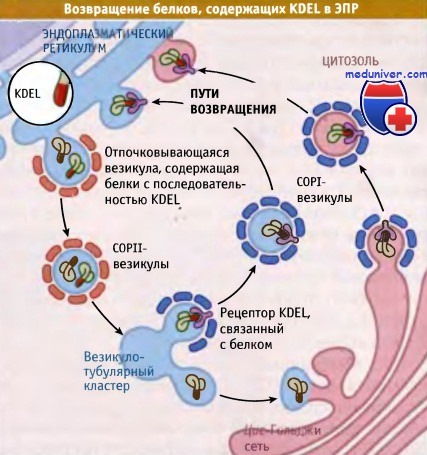
Как следует из рисунка ниже, белки узнаются посредством рецептора, который связывается с сигналом возвращения. В клетках млекопитающих рецепторы KDEL сосредоточены, главным образом, в пост-ЭПР компартментах VTC (везикуло-тубулярных кластерах) и CGN (цис-Гольджи-сети), однако небольшое их количество находится в более дистальных компартментах аппарата Гольджи. Связывание рецептора KDEL с соответствующей последовательностью белка приводит к включению этого комплекса в окаймленные везикулы, называемые COPI, посредством которых комплекс возвращается назад, в ЭПР.
Там происходит диссоциация белка от рецептора. Этот процесс, возможно, инициируется повышенной концентрацией ионов кальция в люмене ЭПР, которая оказывается выше, чем в VTC и CGN. In vitro диссоциация происходит при изменении pH, однако неизвестно, насколько разные величины pH имеют ЭПР и CGN. Ненагруженные рецепторы KDEL выходят из ЭПР в везикулах COPII, направляясь в VTC и CGN, для следующих циклов возвращения.
Из мембран эндоплазматического ретикулума (ЭПР) также могут выходить резидентные белки, которые тоже содержат в своей структуре сигналы возвращения. Эти сигналы состоят из последовательности остатков диаминомонокарбоновых кислот. Резидентные трансмембранные белки, относящиеся к типу I (N-терминальный участок которых находится со стороны люмена), содержат сигнал, включающий два лизиновых остатка (Лиз-Х-Лиз-Х-Х, где X — любая аминокислота), расположенный на конце цитоплазматических участков.
Этот сигнал непосредственно связывается с а-субъединицей белка окаймления COPI. Резидентные ЭПР белки, относящиеся к типу II мембранных белков (С-терминальный участок которых находится со стороны люмена), содержат другой сигнал возврата, для которого характерно наличие двух расположенных рядом остатков аргинина.
Для мембранных белков, которые обычно перемещаются между ЭПР и аппаратом Гольджи, также характерен сигнал возвращения, состоящий из основных аминокислот. В качестве примера приведем ERGIC-53, который транспортирует растворимые белки от ЭПР в аппарат Гольджи. ERGIC-53 имеет как сигнал экспорта из ЭПР, так и сигнал возвращения в ЭПР, содержащий два остатка лизина и расположенный со стороны цитоплазмы.
Белки, присутствующие в люмене ЭПР в высокой концентрации,
иногда выходят в везикулярно-тубулярные кластеры и в цис-компартмент Гольджи.
В структуре этих белков присутствует KDEL последовательность,
узнаваемая KDEL-рецептором, который возвращает их в ЭПР.
Возвращение резидентных растворимых и трансмембранных белков
в эндоплазматический ретикулум происходит с участием специфических сигналов.