• Для удержания белков в аппарате Гольджи достаточно домена, пронизывающего мембрану, и фланкирующих его последовательностей
• Механизм удержания белков Гольджи зависит от их способности образовывать олигомерные комплексы и от длины домена, пронизывающего мембрану
До сих пор мы рассматривали сигналы сортировки, которые обеспечивали селективный транспорт карго из одного компартмента в другой при экзоцитозе в прямом или обратном направлении. Однако существует другой важный механизм, который обеспечивает адресную локализацию индивидуальных белков. Это механизм селективного удержания. Он позволяет определенному белку закрепляться на достигнутом месте и предотвращает его дальнейший неизбирательный или сигнал-зависимый транспорт.
Аппарат Гольджи является одним из мест в клетке, где реализуется механизм селективного удержания белков.
В аппарате Гольджи содержится много ферментов, которые модифицируют олигосахаридные цепи. Их эффект в отношении N-олигосахаридов показан на рисунке ниже. В различных участках стопки цистерн (цис-, транс- или в середине) могут локализоваться различные ферменты. В общем, ферменты Гольджи относятся к трансмембранным белкам типа II с коротким N-терминальным участком, обращенным в сторону цитоплазмы, и с С-терминальным, каталитическим доменом, обращенным в люмен.
Домен, пронизывающий мембрану, представляет собой сигнальный якорь, который сначала направляет вновь образованный фермент Гольджи в ЭПР. Аминокислоты, расположенные по бокам этого домена, обеспечивают надлежащую топологию фермента в мембране. Фермент удерживается в аппарате Гольджи, главным образом, за счет домена и фланкирующих его аминокислот. Если к белку-репортеру, который обычно не локализован в аппарате Гольджи, присоединить пронизывающий мембрану домен и фланкирующие аминокислоты, то этого оказывается достаточно для того, чтобы белок остался локализованным в аппарате Гольджи. Поэтому эти последовательности называются сигналы удержания.
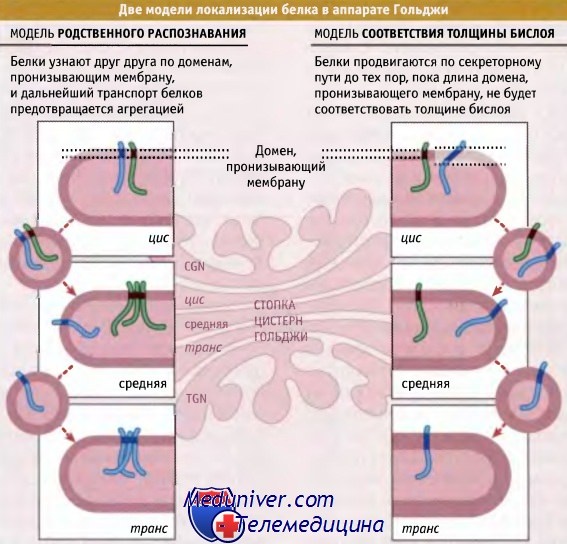
Предложено две модели, объясняющие удержание мембранных белков в аппарате Гольджи: модель родственного распознавания и модель соответствия толщины бислоя. Модель родственного распознавания предполагает, что ферменты, расположенные в одной и той же цистерне аппарата Гольджи, узнают друг друга по своим доменам, пронизывающим мембрану. Если концентрация ферментов достигает некоего критического уровня, то при узнавании образуются настолько большие агрегаты, что они не могут поместиться в COPI-везикулы. Поэтому ферменты не транспортируются ни в прямом, ни в обратном направлении.
Согласно этой модели, удержание белка в аппарате Гольджи не связано с наличием специальных рецепторов (в отличие от возвращения белков, обусловленного, например, рецептором KDEL). Таким образом, удержание белков в аппарате Гольджи по данной модели представляет собой процесс, который не характеризуется насыщением.
Модель родственного распознавания поддерживается результатами экспериментов по искусственному удержанию ферментов Гольджи в ЭПР. При этом остальные ферменты, обычно расположенные в тех же цистернах, перемещаются в другие места. В этих экспериментах в клетке гиперэкспрессировался фермент Гольджи, который в цитоплазматическом домене содержал сигнал удержания в ЭПР. Молекулы такого мутантного фермента были обнаружены в ЭПР в составе комплекса с молекулами фермента дикого типа. Таким образом, под влиянием мутантного фермента его дикая форма переместилась из аппарата Гольджи в ЭПР.
Модель соответствия толщины бислоя предполагает, что в удержании белка в аппарате Гольджи важную роль играет размер домена, пронизывающего мембрану, а не его аминокислотный состав. В основе модели лежит факт увеличения содержания холестерина в мембранах по пути экзоцитоза от места его синтеза в мембране ЭПР к плазматической мембране. Поскольку in vitro увеличение содержания холестерина ведет к увеличению толщины липидного бислоя, ожидается, что в направлении от ЭПР к плазматической мембране должен существовать градиент его толщины.
По сравнению с ферментами плазматических мембран, ферменты аппарата Гольджи обладают более короткими мембранными доменами, поэтому они будут транспортироваться до того момента, пока размер домена не станет соответствовать толщине бислоя. Для дальнейшего транспорта белка необходимо включение заряженных фланкирующих аминокислот в гидрофобный бислой, что затруднено энергетически.
Модель соответствия толщины бислоя подтверждается экспериментами, в которых в одном из ферментов Гольджи домен, пронизывающий мембрану, был замещен последовательностью, состоящей из нескольких остатков лейцина, который представляет собой гидрофобную аминокислоту, соответствующую окружению внутри липидного бислоя. Фермент, содержащий короткую последовательность лейциновых остатков, удерживался в аппарате Гольджи, в то время как фермент с более длинным доменом транспортировался в плазматическую мембрану.
Следует отметить, что предложенные модели не исключают друг друга, и обе могут принимать участие в локализации ферментов в индивидуальных цистернах аппарата Гольджи. Агрегированные ферменты Гольджи выходят из цистерн с меньшей вероятностью, поскольку энергетические затраты при несовпадении домена, пронизывающего мембрану, и толщины бислоя в этом случае оказываются гораздо выше, чем для индивидуальных ферментов.
Две модели, описывающие возвращение трансмембранных белков в аппарат Гольджи:
модель родственного распознавания и модель соответствия толщины бислоя.