Главный комплекс гистосовместимости (HLA): структура, гены
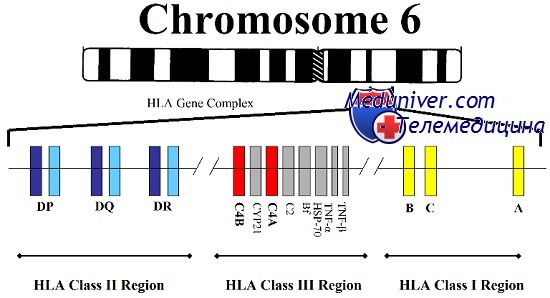
МНС формируется большой группой генов, расположенных на коротком плече хромосомы 6. На основе структурных и функциональных различий эти гены подразделяют на три класса, два из которых, класс I и класс II, относятся к генам HLA, первоначально обнаруженных благодаря их значению в пересадке тканей между неродственными индивидуумами.
Гены HLA класса I и II кодируют поверхностные белки клеток, играющие определяющую роль в инициации иммунного ответа, особенно в «распознавании» антигена лимфоцитами, которые не могут реагировать на антиген, если он не образует комплекс с молекулой HLA на поверхности содержащей антиген клетки. Известно много сотен разных аллелей HLA класса I и И, и ежедневно обнаруживают новые аллели, что делает их наиболее полиморфными локусами в геноме человека.
Гены класса I (HLА-А, HLA-B и HLA-C) кодируют белки, выступающие неотъемлемой частью плазматической мембраны всех ядерных клеток. Белки класса I состоят из двух полипептидных субъединиц: вариабельной тяжелой цепи, кодируемой в пределах МНС, и неполиморфного полипептида, b2-микроглобулина, кодируемого геном, расположенным за пределами МНС и картированным на хромосоме 15. Производные от внутриклеточных белков пептиды образуются путем протеолитического расщепления большими многофункциональными протеазами; затем пептиды перемещаются на поверхность клетки и прикрепляются к молекулам класса I, формируя пептидный антиген для цитотоксических Т-клеток.
Регион класса II состоит из нескольких локусов, таких как HLA-DP, HLA-DQ и HLA-DR, кодирующих поверхностные белки клеточной оболочки. Каждая молекула класса II — гетеродимер, сформированный из а- и b-субъединиц, закодированных в МНС. Молекулы класса II представляют пептиды, производные от внеклеточных белков, которые захватываются лизосомами и перерабатываются в пептиды, узнаваемые Т-клетками.
В пределах МНС присутствуют локусы и других генов, но не имеющих функционального отношения к генам HLA класса I и II и не определяющих гистосовместимость или иммунные ответные реакции. Некоторые из этих генов, тем не менее, связаны с болезнями, например врожденной гиперплазией надпочечников, вызываемую недостаточностью 21-гидроксилазы, и гемохроматозом, болезнью печени, вызванной накоплением железа.
Аллели и гаплотипы главного комплекса гистосовместимости (HLA)
Система HLA сначала может показаться запутанной, поскольку номенклатура, используемая для определения и описания разных аллелей HLA, подвергалась фундаментальным изменениям по мере распространения секвенирования ДНК МНС. Согласно более старой, традиционной системе номенклатуры HLA, разные аллели отличались друг от друга серологически. Индивидуальные типы HLA определялись тем, как панель различных антисывороток или чувствительных лимфоцитов реагирует на клетки.
Антисыворотки и клетки получали от сотен беременных, развивших иммунный ответ против отцовских антигенов I и II типа, экспремируемых плодами в ходе беременности. Если клетки от двух не связанных родственными отношения индивидуумов вызывали ту же реакцию при внесении в панель антител и клеток, считали, что они имеют те же типы и аллели HLA, обозначаемые их номером, например В27 в локусе HLA-B класса I или DR3 в локусе DR класса II.
Тем не менее после идентификации и секвенирования генов, ответственных за кодирование цепей МНС класса I и класса II, отдельные первоначально определенные серологически аллели HLA даже в пределах одного серологического аллеля оказались состоящими из многочисленных аллелей, определяемых различными вариантами последовательности ДНК. 100 серологически определяемых типов HLA-A, В, С, DR, DQ и DP теперь включают более 1300 аллелей, определяемых на уровне последовательности ДНК.
Например, в гене HLA-B, прежде определявшемся серологической реакцией как единый аллель В27, обнаружено более 24 различных вариантов последовательности нуклеиновых кислот. Большинство, хотя и не все, варианты ДНК представляют изменение кодона триплета и, следовательно, аминокислоты в пептиде, кодируемом этим аллелем. Каждый аллель, изменяющий аминокислоту в пептиде HLA-B, получает свой дополнительный последовательный номер, например аллель1, 2, и так далее в группе аллелей, соответствующих ранее единственному аллелю В27, и теперь называется HLA-B*2701, HLA-B*2702 и так далее.
Набор аллелей HLA в различных локусах класса I и II в данной хромосоме формирует гаплотип. Аллели кодоминантны; каждый из родителей имеет два гаплотипа и экспрессирует их оба. Эти локусы располагаются достаточно близко друг к другу, так что в конкретной семье гаплотип может передаваться ребенку как единый блок. В результате родитель и ребенок имеют общий гаплотип, а шанс, что два сибса унаследуют один гаплотип HLA, равен 25%.
Поскольку приживление пересаженных тканей в основном согласуется со степенью сходства между HLA гаплотипами донора и реципиента (и группы крови АВО), лучший донор костного мозга или органа — АВО-совместимый и HLA-идентичный сибс реципиента.
В любой этнической группе некоторые аллели HLA обнаруживаются часто, а другие — редко или никогда. Аналогично некоторые гаплотипы встречаются чаще, чем ожидалось, тогда как другие — исключительно редко или не встречаются вовсе. Например, большинство из Зх107 теоретически возможных комбинаций аллелей в гаплотипе в белой популяции никогда не встречаются. Это ограничение в разнообразии гаплотипов в популяции, возможно, вызвано ситуацией, получившей название неравновесного сцепления и может объясняться сложным взаимодействием множества факторов.
Эти факторы включают низкие показатели мейотической рекомбинации из-за небольшого расстояния между локусами HLA; влияние окружающей среды, обеспечивающее положительный отбор для конкретных комбинаций аллелей HLA, формирующих гаплотип; и исторические факторы, например, как давно образовалась популяция, число основателей и интенсивность происходившей иммиграции (см. далее в этой главе).
Между популяциями также существуют значительные различия в частоте аллелей и гаплотипов. То, что бывает частым аллелем или гаплотипом в одной популяции, может оказаться очень редким в другой. Различия в распределении и частоте аллелей и гаплотипов в пределах МНС — результат сложного взаимодействия генетических, средовых и исторических факторов в каждой конкретной популяции.