Факторы стимулирующие рост клетки и аутокринный цикл
• Для функционирования сигнальной системы клетке необходимы внеклеточные факторы, рецепторы и другие белки, передающие сигнал в ядро
• Сигналы, поступающие в клетку, могут активировать или ингибировать ее рост
• Многие гены, кодирующие сигнальные молекулы клетки, представляют собой протоонкогены и гены-супрессоры опухоли
Для того чтобы понять, каким образом мутации в онкогенах и генах-супрессорах опухоли влияют на рост и пролиферацию клеток, мы вначале должны рассмотреть, как регулируются эти процессы в здоровых клетках. Клетки реагируют на разнообразные поступающие к ним сигналы, и в особенности на доставляемые ростовыми факторами. Обычно эти факторы высвобождаются определенными клетками в ткани, проходят через межклеточное пространство, и попадают в клетку-мишень, в которой в ответ происходит активация или ингибирование пролиферации.
Способность клетки-мишени реагировать противоположным образом на сигнал предполагает наличие сложного клеточного механизма, обеспечивающего получение внешнего сигнала, его обработку и исполнение решения об изменении скорости роста клетки. В качестве примера функционирования клеточных систем передачи сигнала и его нарушения мы рассмотрим внутриклеточные сигналы, которые генерируются Ras и TGF-b.
На поверхности клетки расположено множество рецепторов, способных узнавать и связывать ростовые факторы, причем каждый рецептор специфически связывается с сооответствующим лигандом. Например, PDGF специфически связывается с рецептором PDGF, расположенным на поверхности клетки, a EGF — с рецептором EGF. Когда рецептор связался с лигандом, он передает сигнал через плазматическую мембрану внутрь клетки.
В свою очередь, этот сигнал проходит по сложному каскаду передающих белков, которые функционируют наподобие молекулярной «пожарной цепочки», каждый член которой получает сигнал от партнера, обрабатывает и усиливает его, и затем передает следующему или следующим партнерам.
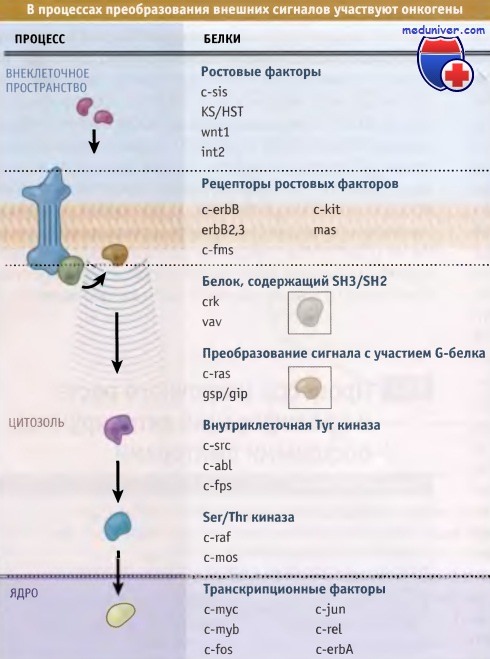
Белки, кодируемые протоонкогеном, можно обнаружить во многих звеньях передачи сигнала.
Многие белки, участвующие в преобразовании ростовых сигналов, могут стать онкобелками.
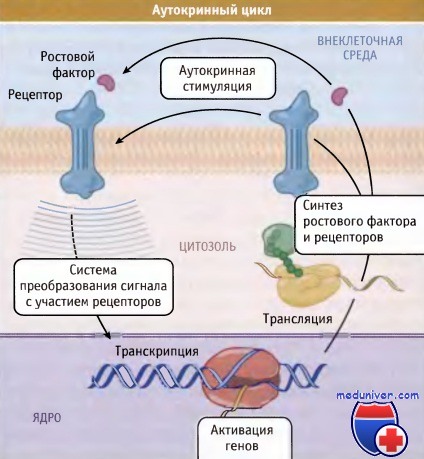
Одна группа протоонкогенов кодирует белки ростового фактора. Если в неподходящее время какой-либо ген из этой группы начал экспрессироваться в клетке, которая также содержит рецепторы для этого фактора, то возникает постоянная положительная обратная связь. Таким образом, клетка высвобождает в окружающую среду большие количества митогенного фактора роста, кодируемого онкогеном. Затем этот ростовой фактор связывается с рецепторами на поверхности этой же клетки и активирует их, что, в свою очередь, стимулирует клетку постоянному росту.
Это приводит к независимости клетки от ростового фактора, поскольку она сама вырабатывает фактор, генерирующий митотические сигналы. Такая самостимуляция роста называется аутокринной регуляцией. Она отличается от паракринной регуляции, при которой соседние клетки обмениваются сигналами, и эндокринной регуляцией, которая зависит от факторов, образующихся в определенных органах и проходящих большие расстояния по системе кровообращения, прежде чем попасть в соответствующую клетку-мишень. В качестве примера приведем онкоген sis, кодирующий одну из форм PDGF, которая образуется постоянно и с высокой скоростью.
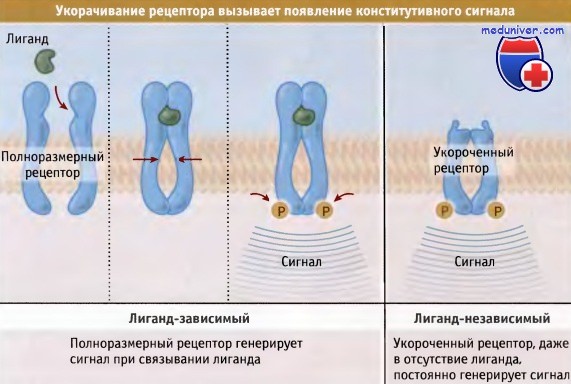
Вторая группа протоонкогенов кодирует рецепторы ростовых факторов. Эти рецепторы при нарушениях функционирования посылают в клетку сигналы стимуляции роста, даже при отсутствии связывания с соответствующими лигандами. В данном случае клетка опять становится независмой от ростовых факторов, поскольку пролиферирует в отсутствие митогенных факторов, обычно необходимых для запуска этого процесса. Фактически существует два механизма нарушения функционирования рецепторов GF, которые кодируются протоонкогенами. На рисунке ниже показано, что некоторые онкогены кодируют транкированные GF-рецепторы.
Во многих опухолях человека, например, обнаруживаются транкированные GF-рецепторы, у которых отсутствует внеклеточный домен; такие транкированные рецепторы, независимо от присутствия лиганда, постоянно передают сигналы. В других опухолях, например расположенных в молочной железе, головном мозгу и в желудке, наблюдается гиперэкспрессия EGF рецепторов, и они присутствуют в раковых клетках в гораздо большем количестве, чем в здоровых. В данном случае снова рецепторы передают сигнал постоянно и независимо от присутствия лиганда.
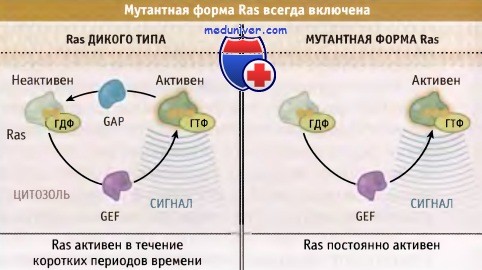
Внутриклеточные белки, участвующие в обработке сигнала, поступающего от рецепторов, также являются мишенями, воздействие на которые вызывает активацию онкогенов. Хорошим примером такого типа мишени является белок, кодируемый протоонкогеном ras. Этот белок обычно находится в цитоплазме в ожидании сигнала, поступающего от рецептора, расположенного на поверхности клетки. Когда рецептор связывается с лигандом и активируется, он посылает сигнал, который поступает к Ras белку, пройдя через несколько промежуточных белков.
Белок Ras реагирует на этот сигнал, превращаясь в активированную форму, способную генерировать сигнал. В таком активном, способном к генерации сигнала состоянии белок существует лишь в течение короткого времени — от нескольких секунд до минуты, после чего выключается, прекращает генерировать сигнал и возвращается в покоящееся, неактивное состояние.
В клетках, несущих мутацию по онкогену ras, образуется белок Ras с измененной структурой, который по-прежнему способен активироваться и генерировать сигнал, но не способен выключаться. Поэтому он может пребывать в таком активном состоянии много минут и даже часов, генерируя в клетке огромное количество сигналов стимуляции роста. Такое состояние резко отличается от обстановки импульсных митотических сигналов, которые генерируются нормальным Ras белком. На рисунке ниже показаны различия между активированным и нормальным Ras.
Подобные белки с нарушенной функцией обнаружены примерно в четверти опухолей человека различного происхождения.
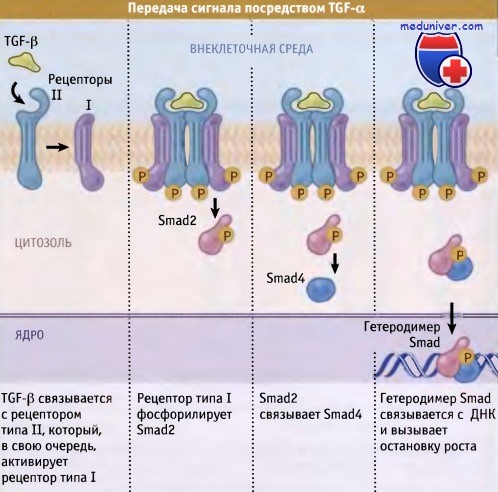
Клетки также могут реагировать на поступающие сигналы ингибированием роста и пролиферации. Как отмечалось ранее, TGF-b представляет собой сигнальную молекулу, обладающую подобным отрицательным эффектом на рост клеток. TGF-b на определенное время переводит клетку в неактивное, покоящееся состояние, из которого, при необходимости, она может в будущем выйти. Раковые клетки становятся устойчивыми (или рефрактерными) к подобным сигналам ингибирования роста таким же образом, как они могут избегнуть необходимости стимуляции ростовым фактором. У некоторых раковых клеток не происходит экспрессия TGF-b рецепторов.
У других прекращается образование одного из критических белков в системе передачи сигнала, принадлежащего к классу белков Smad. Эти белки участвуют в передаче сигнала от TGF-b рецепторов в ядро клетки. В некоторых случаях образуются дефектные Smad белки, которые не способны правильно передавать сигналы от TGF-b рецепторов. Иногда в клетках отсутствуют компоненты ядерного механизма, которые позволяют им прекращать пролиферацию в ответ на поступление сигнала от рецепторов TGF-p. Здесь мы имеем дело с ситуацией, характер которой как бы напоминает зеркальное отражение по отношению к митогенным сигналам. В отличие от усиления сигнала белками-стимуляторами роста, в этих случаях белки-супрессоры опухоли, ответственные за ингибирование пролиферации клеток, инактивируются.
Когда клетка продуцирует собственный ростовой фактор,
возникает самоподдерживающийся аутокринный цикл.
В некоторых случаях удаление из рецептора домена связывания лиганда приводит к появлению конститутивного сигнала,
поскольку для димеризации рецептора больше не требуется связывания лиганда.
Некоторые мутантные формы Ras постоянно находятся в конформации,
содержащей связанный ГТФ, и постоянно поддерживают в активном состоянии внутриклеточную систему преобразования внешнего сигнала.
TGF-b служит одним из сигналов подавления роста, которые получает клетка.
После получения этого сигнала в клетке запускается сигнальный каскад,
в результате чего активируются Smad белки, которые активируют транскрипцию белков, подавляющих рост.