В норме движения глаз синхронны, что формирует два принципиально различных вида движения:
1. Перемещение взора. Данные произвольные движения глаз регулируют корковые и подкорковые структуры.
а. Саккадные движения глаз, или быстрые однонаправленные движения глаз, с помощью которых происходят перенаправление взора на новый объект интереса и фиксация изображения на центральной ямке.
б. Вергентные движения глаз, или разнонаправленные движения глаз, которые регулируют фиксацию взора от далеко расположенных объектов на близко расположенные (например, фиксационный рефлекс).
2. Фиксация взора. Зрительная система (от сетчатки до зрительной коры) контролирует обратную связь относительно положения рассматриваемой цели и положения глаз, которая обеспечивает фокусировку объекта на сетчатке глаза при перемещении объекта или индивида в пространстве.
а. Слежение, или плавные движения, происходит, когда глаза наблюдают за объектом интереса при его медленном передвижении в поле зрения. При этом скорость передвижения глаз совпадает с наблюдаемой скоростью, и сохраняется фокусировка изображения на центральной ямке. (В отсутствие объекта для отслеживания человек не может сознательно перемещать взгляд с такой низкой скоростью. Попытки сделать это приводят к небольшим саккадным движениям глаз.)
б. Вестибулоокулярный рефлекс. С помощью данного рефлекса можно удерживать взгляд на предмете интереса во время движений головы. Он формируется при перемещении эндолимфы в кинетическом лабиринте.
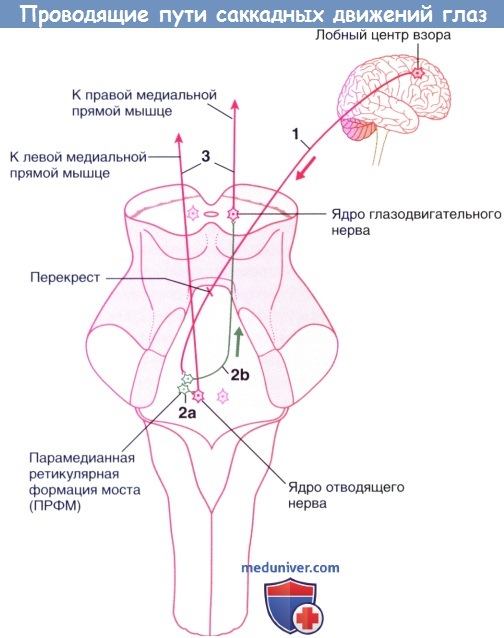
Проводящие пути, участвующие в формировании произвольных саккандных движений глаз влево.
(1) Аксоны от правого лобного центра взора активируют парамедианную ретикулярную формацию моста (ПРФМ).
(2) Некоторые нейроны ПРФМ активируют прилежащие нейроны отводящего ядра (3).
Другие нейроны ПРФМ отправляют обильно миелинизированные (быстрые) межъядерные волокна по медиальному продольному пучку для активации глазодвигательных нейронов, обеспечивающих активность правой медиальной прямой мышцы.
Синхронные сокращения соответствующих прямых мышц образуют саккадные движения влево.
а) Перемещение взора. Четыре самостоятельных центра взора регулируют активность моторных нейронов в соответствии с направлением движения (влево, вправо, вверх или вниз) и участвуют как в перемещении взора, так и в его фиксации. Центры взора представляют собой небольшие узелки в ретикулярной формации. Они содержат нейроны с высокой лабильностью (1000 Гц, или импульсов/с), которые мгновенно активируют соответствующие моторные нейроны с большой скоростью, поскольку необходимо преодолеть упругие свойства орбиты для инициации движения глаз.
Парные центры (слева и справа) для горизонтальных движений глаз расположены в парамедианной ретикулярной формации моста (ПРФМ). При активации каждого центра происходят сопряженные движения глаз в свою сторону. Например, при активации левой ПРФМ происходит перемещение линии взора обоих глаз влево. На уровне среднего мозга находится билатеральный центр, регулирующий вертикальные движения глаз. Он расположен на ростральном конце медиального продольного пучка (МПП) на уровне предкрышечного ядра — рострального интерстициального ядра МПП (риМПП). Вероятно, нейроны данного ядра для взгляда вверх расположены кзади, а для взгляда вниз — впереди. (Другие ядра, участвующие в регуляции вертикальных движений глаз, включают интерстициальное ядро Кахаля (ИЯК), которое расположено вентральнее на том же уровне и играет определенную роль в интеграции информации из продолговатого мозга и моста, а также ядро Даркшевича, которое способствует формированию взгляда вверх и соответствующих движений век.)
Автоматические сканирующие движения активируются через верхний бугорок при получении визуальной информации от сетчатки через зрительный тракт.
Примером автоматических сканирующих движений может служить быстрый взгляд в сторону объекта внимания на периферии поля зрения. Соответствующие покрышечно-ретикулярные волокна пересекают среднюю линию перед вступлением в центр взора.
Точные саккадные движения контролирует червь мозжечка, к которому подходят афференты от верхних бугорков четверохолмия. От червя мозжечка проводящий путь направляется к вестибулярному ядру.
Задняя теменная кора головного мозга, вероятно, участвует в перемещении взора на новые объекты внимания, а поле взора теменной коры больших полушарий (поле Бродмана 7а) активируется при анализе визуального образа при помощи контролируемых саккадных движений. Обе области активно взаимодействуют с полем взора лобной коры больших полушарий.
Произвольные саккадные движения глаз в большей степени изучены при горизонтальных движениях глаз. При этом существует несколько областей в лобной коре больших полушарий (фронтальное поле взора отвечает за произвольные саккады, а также саккады, связанные с долговременной памятью, и вергентные движения; дополнительное поле взора—за планирование и запоминание саккадных движений глаз; дорсолатеральная префронтальная кора—за выполнение спланированных саккадных движений к запомненным целям), которые двунаправленно взаимодействуют с теменной корой для выполнения подобных произвольных движений глаз. Аксоны от лобной коры головного мозга спускаются к переднему бедру внутренней капсулы, и большинство из них переходит на другую сторону перед вступлением в противоположный центр взора ствола головного мозга. Другие аксоны от лобной коры головного мозга оканчиваются синапсами на базальных ганглиях (хвостатое ядро, черная субстанция), которые помогают поддерживать «баланс» между рефлекторными и произвольными движениями для предотвращения нежелательных саккад. Их действие реализуется посредством группы нейронов, расположенных диффузно в среднем мозге и мозжечке (универсальные паузовые нейроны). В то же время, как будет отмечено в отдельной статье на сайте, верхние холмики четверохолмия противоположной стороны активируются для усиления возбуждения в соответствующем центре взора.
Контроль горизонтальных саккадных движений осуществляет полушарие противоположной стороны: левое полушарие отвечает за саккадные движения глаз направо и наоборот. У некоторых пациентов при остром нарушении кровообращения в области фронтальной коры (в большинстве случаев — при инсульте) можно временно наблюдать невозможность произвольного взгляда в сторону, противоположную повреждению (паралич взора, или приобретенная глазодвигательная апраксия), в то время как вестибулоокулярный рефлекс остается неповрежденным. Подобный паралич взора обычно проходит в течение недели, даже при остаточной выраженной гемиплегии. Односторонние поражения коры теменной доли, особенно с правой стороны, могут привести к замедленным или гипометричным (с меньшей амплитудой) саккадным движениям на стороне, противоположной поражению.
б) Фиксация взора. Нейронные механизмы, обеспечивающие слежение, обладают сложной организацией и включают: (а) целостность зрительных путей для наблюдения за расположением объекта во время его движения; (б) нейроны, обеспечивающие информацию о скорости движения объекта (детекторы скорости); (в) нейроны, обеспечивающие координацию движений глаз и головы (нейронный интегратор); (г) систему контроля плавного выполнения следящего перемещения. Примером работы подобной системы плавного слежения за объектом в горизонтальной плоскости может служить следующее.
• Регистрация движущегося объекта начинается в сетчатке, распространяется по зрительному нерву к латеральным ядрам коленчатого тела и далее — к первичной зрительной коре. Затем происходит слияние первичного аксона с аксонами лобной, теменной и височной коры в области височно-теменно-затылочного перехода, откуда аксоны направляются по той же стороне к мосту.
• От ипсилатерального дорсолатерального ядра моста аксоны направляются к мозжечковому клочку противоположной стороны, а затем — к вестибулярному ядру.
• От вестибулярных ядер аксоны направляются назад (на той же стороне, что и изначальный корковый ответ) к мосту и ПРФМ, вследствие чего формируется конечный результат — сопряженное горизонтальное движение глаз. Необходимо помнить, что плавные следящие движения обеспечивают структуры той же стороны, что и направление движений глаз: правое полушарие отвечает за плавное следящее движение глаз вправо и наоборот.
Вертикальные движения глаз обеспечивает активность правого и левого полушарий головного мозга. Аксоны от коры полушарий идут к риМПП, откуда они направляются к соответствующим двигательным нейронам III и IV пар черепных нервов. ИЯК интегрирует дополнительную информацию от нейронов вестибулярного аппарата, моста и продолговатого мозга, а затем ядро Даркшевича регулирует движения глаз вверх и движения верхнего века.
Вестибулоокулярный рефлекс формируется динамической частью лабиринта и обеспечивает интеграцию пространственной и скоростной информации об объекте от предподьязычного ядра — узла ретикулярной формации, который на самом деле расположен ближе к ядру отводящего нерва, чем к ядру подъязычного нерва. От данного ядра аксоны направляются к ПРФМ или (для вертикальных движений глаз) к риМПП для формирования сопряженных движений глаз. Аксоны от коры больших полушарий (фронтальная, теменная и кора островковой доли) дополнительно регулируют движения глаз. Корковая регуляция в качестве рефлекторной реакции также должна быть подавлена в случае, если цель и голова (и глаза) движутся синхронно в одном направлении. В противном случае рефлекс будет формировать движения глаза в направлении, противоположном направлению движения головы!
Совместная деятельность динамического лабиринта и мозжечка обеспечивает фиксацию глаза на цели во время движения головы.
в) Резюме:
1. Глазодвигательный нерв. Соматические эфферентные волокна III пары черепных нервов берут начало от основного ядра на уровне верхних холмиков четверохолмия. Нерв проходит через пещеристый синус и разделяется на две ветви после вступления в верхнюю глазничную щель. Верхняя ветвь иннервирует верхнюю прямую мышцу и мышцу, поднимающую верхнее веко, а нижняя ветвь—нижнюю и медиальную прямые мышцы, а также нижнюю косую мышцу.
Парасимпатические волокна выходят из ядра Эдингера-Вестфаля, идут в сопровождении основного нерва и образуют синапсы в ресничном ганглии, иннервирующем сфинктер зрачка и ресничную мышцу.
Паралич глазодвигательного нерва проявляется расширением зрачка, птозом верхнего века, а затем расходящимся косоглазием.
2. Блоковый нерв. Ядро IV пары черепных нервов находится на уровне нижних холмиков четверохолмия. Нерв пересекает среднюю линию до выхода ниже уровня нижнего холмика, проходит через пещеристый синус и иннервирует верхнюю косую мышцу.
Паралич данного нерва характеризуется диплопией при взгляде вниз. 3. Отводящий нерв. Ядро находится на уровне лицевого бугорка в мосту. Нерв идет в субарахноидальном пространстве от нижней границы моста к верхушке каменистой части височной кости, проходит через пещеристый синус, верхнюю глазничную щель и иннервирует латеральную прямую мышцу.
Паралич данного нерва характеризуется сходящимся косоглазием с невозможностью отведения в пораженном глазу. Существует простое мнемоническое правило для запоминания иннервации глазодвигательных мышц: ЛП6(ВК4)3. Эту «химическую формулу» расшифровывают следующим образом: латеральную прямую мышцу (ЛП) иннервирует VI пара черепных нервов, верхнюю косую мышцу (ВК) иннервирует IV пара черепных нервов, а остальную часть глазных мышц иннервирует III пара черепных нервов.
4. Симпатическая иннервация. Симпатическая иннервация (через α1-арецепторы) стимулирует дилататор зрачка и гладкую мышцу в переднем отделе мышцы, поднимающей верхнее веко (верхняя тарзальная мышца Мюллера). Паралич данного нерва характеризуется птозом и сужением зрачка (синдром Горнера). Симпатическая иннервация (через β2-ре-цепторы) подавляет активность сфинктера зрачка и ресничной мышцы. 5. Парасимпатическая иннервация. Парасимпатическая иннервация стимулирует сфинктер зрачка и ресничную мышцу. 6. Проводящие пути. Для рефлекторного сужения зрачка на свет: от сетчатки к предкрышечным ядрам, к обоим ядрам Эдингера-Вестфаля, затем к ресничному ганглию и сфинктеру зрачка.
Для аккомодационного рефлекса: от сетчатки к боковым коленчатым телам, к затылочной коре, к ядрам Эдингера-Вестфаля, к ресничному ганглию и ресничной мышце.
7. Контроль движений глаз. Перемещение взора (саккадные движения) локально активируют шесть центров взора. Наиболее важный с клинической точки зрения—ПРФМ, которая регулирует работу латеральной прямой мышцы на той же стороне и сопряженные движения медиальной прямой мышцы на своей стороне. Автоматические сканирующие движения контролируют верхние холмики четверохолмия, а произвольные—поля зрения фронтальной коры полушарий головного мозга.
Фиксацию взора обеспечивает ряд структур, среди которых: затылочная кора головного мозга, динамическая часть лабиринта, мозжечок, верхние холмики четверохолмия и ретикулярная формация.