Легкие цепи иммуноглобулинов. Организация иммуноглобулинов
V- и С-области легких цепей примерно равны по длине. Так, у каппа-цепей человека V-области строятся из 107—113 аминокислотных участков (начиная с N-конца), а на С-область приходятся остальные 107 остатков. У тяжелых гамма-цепей длина V-областей примерно такая же, а С-область состоит из трех линейно расположенных друг за другом гомологичных участков С1, С2, С3 сходной длины, структурно гомологичных с С-областями легких цепей и друг с другом.
V-области легких каппа- и ламбда-цепей различаются между собой, и они иные, чем у тяжелый цепей, тогда как V-области тяжелых цепей одинаковы у всех классов.
Среди легких цепей, выделенных из разных патологических иммуноглобулинов, нет ни одной, похожей на другие. Подсчитано, что различных идиотипических вариантов существует не менее нескольких тысяч. Несмотря на это многообразие, все варианты V-областей каппа-цепей, укладываются в три основные подгруппы, причем различия между белками каждой подгруппы невелики.
Таким же путем удалось классифицировать V-области ламбда-цепей, а в последнее время и V-области тяжелых цепей.
Аминокислотные замены, обусловливающие различия между V-областями, в большинстве своем сгруппированы в нескольких определенных участках — так называемых горячих точках. У легких цепей эти гипервариабельные области занимают положения 25—35, 52—55 и 89—96, а у тяжелых гамма-цепей — 31—37, 86—91 и 101 —109. Эти остатки, очевидно, участвуют в формировании активного центра антителВесьма закономерно в пептидных цепях иммуноглобулинов расположение внутрицепьевых дисульфидных мостиков.
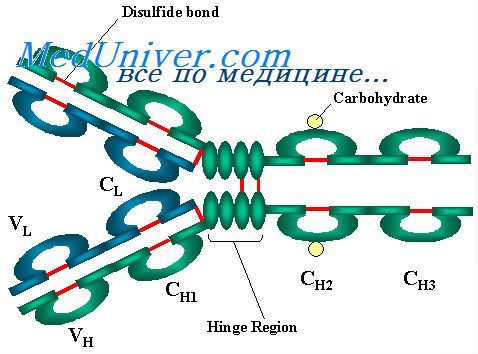
Каждый из них образует петлю из части цепи длиной примерно в 60 остатков, а вправо и влево от нее имеется но 20 остатков. Таким образом, основной структурной единицей цепей является участок примерно в 100 остатков с одним дисульфидным мостиком. В легких цепях их два и они соответствуют V- и С-областям, а в гамма-цепях их четыре. В нативной цепи каждый из этих участков свернут в отдельную, относительно независимую компактную глобулу-домен (Putman, 1969).
Сейчас можно считать доказанной схему строения молекул IgG, предложенную Портером (Porter, 1973). Согласно этой схеме, молекула строится из L- и Н-цепей, соединенных мсжцепьевыми ди-сульфидными мостиками. При протеолизе (например, папаино.м или трипсином), как показал Портер, молекула распадается на три фрагмента, два из которых идентичны друг другу и обладают антительной активностью (Fab-фрагменты). Они состоят из легкой цепи и половины тяжелой цепи. Третий фрагмент (Fc) состоит из С-концевых половин тяжелых цепей.
Число связей между тяжелыми цепями варьирует и у разных подклассов IgG человека составляет от двух до пяти.
IgA встречается в двух формах — мономерной, сходной с молекулой IgG, и полимерной, строящейся из двух или более мономерных единиц. Макроглобулины IgM построены из пяти мономерных субъединиц, напоминающих молекулу IgG.
Между разными по специфичности антителами, принадлежащими к одному классу (подклассу) и типу, крайне трудно обнаружить какие-либо различия в свойствах, за исключением их способности реагировать с антигенами. Таким образом, структура антител к самым разным антигенам весьма сходная, за исключением того небольшого участка, которым они непосредственно контактируют с антигенными детерминантами.
Антигенами же могут быть самые разнообразные вещества (белки, полисахариды, синтетические вещества), которые имеют сильно отличающуюся конфигурацию и, по всей видимости, весьма различным образом расположенные антигенные детерминанты.