Как известно (Wu, Kabat, 1970), по первичной структуре остатки вариабельной области можно разделить на гипервариабельные, которыми обусловлены основные различия между цепями, и нсгипервариабельные, или каркасные, остатки. Гипервариабельные остатки доступны растворителям и у различных вариабельных доменов свернуты различным образом.
Так, например, первая гипервариабельная область легких цепей (L1) у белка Rei представляет собой вытянутую цепь, тогда как у белков New и Meg она свернута в спираль. В этом месте у белка МсРС 603 имеется вставка из шести остатков аминокислот, и эта область представляет собой вытянутую неспиральную петлю. В противоположность зтому, хотя и с небольшими вариациями, расположение каркасных, неварьирующих остатков у разных доменов очень сходное.
Эта гомология в третичной структуре вариабельных доменов указывает на то, что неварьирующие остатки создают ригидный каркас, в который как бы встраиваются гипервариабельные петли.
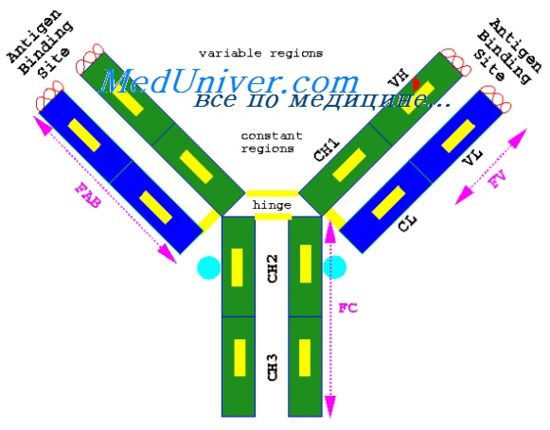
Четыре домена (два вариабельных и два постоянных) образуют один Fab-фрагмент. При этом отмечаются следующие закономерности.
1. Оба постоянных домена взаимодействуют более тесно, что обусловливает большую компактность всей постоянной области по сравнению с вариабельной. Это объясняется тем, что постоянные домены взаимодействуют между собой четырехсегментными слоями, образуя большую контактирующую поверхность, тогда как вариабельные домены взаимодействуют трехсегментными слоями.
2. Основные взаимодействия между доменами являются латеральными, т. е. и вариабельные, и постоянные домены взаимодействуют между собой, тогда как продольные взаимодействия (между вариабельными и постоянными) выражены меньше.
3. Оба домена тяжелой цепи, т. е. VH и Сн, находятся в более тесном контакте, чем оба домена легкой цепи, что 'вызывает небольшой изгиб всего Fab-фрагмента. Такой же изгиб присущ димеру легких цепей. Последний строится из двух идентичных по своей первичной -структуре легких цепей — мономеров. Несмотря на это, угол между доменами у одного мономера равен 70°, тогда как у другого — 110°, что и обусловливает изгиб всей молекулы.
Структура Fc-фрагмента отличается отсутствием такого изгиба. Оба СнЗ-домена тесно прилегают друг к другу, подобно CL- и Сн1 -доменам в Fab-фрагменте, тогда как Сн2-домены не контактируют между собой, за исключением ковалентной связи в шарнирной области. Возможно, такое расположение Сн2-доменов необходимо для их взаимодействия с первым компонентом комплемента. Следует отметить, что результаты рентгеноструктурного анализа иммуноглобулинов полностью согласуются с полученными ранее данными других методов.
Так, например, с помощью оптических методов — дисперсия оптического вращения (Троицкий и др., 1971) и инфракрасная спектроскопия водородно-дейтериевым обменом (Abaturov е. а., 1969) — было найдено, что основным элементом вторичной структуры пептидных цепей иммуноглобулинов является бета-складчатость.
Кроме того, есть данные свидетельствующие в пользу гипотезы доменов (Edelman, 1973). Согласно этой гипотезе, каждый гомологичный участок полипептидной цепи иммуноглобулинов длиной около 110 аминокислотных остатков с одной внутренней дисульфидной связью свертывается в отдельную, относительно независимую глобулу — домен. Например, легкие цепи состоят из двух доменов, тяжелые гамма-цепи — из четырех.
Между глобулами имеются открытые участки полипептидной цепи, особенно чувствительные к протеолитическим ферментам. Одним (из самых важных доказательств доменной структуры пептидных цепей иммуноглобулинов как раз и явилась возможность использования протсаз для расщепления легких цепей на половины, которые не изменяли существенно своей третичной структуры по сравнению с таковой в нативной цепи. Последнее было доказано как оптическими методами, так и исследованиями антигенных свойств половин.
Уже к середине 60-х годов были получены весьма обоснованные данные, базирующиеся в основном на изучении рекомбинации тяжелых и легких цепей и мечении по сродству, о том, что аминокислотные остатки как легкой, так и тяжелой цепи участвуют в образовании активного центра. На моделях, основанных на данных рентгеноструктурпых исследований активных Fab-фрагментов, действительно видно, что обе цепи формируют полость активного центра.
Классические исследования Кабата (Kabat, 1976), Каруша (Каrush, 1962) и Села (Sela, 1970) о размерах активного центра также находятся в хорошем согласии с рентгеноструктурными исследованиями фрагментов двух миеломных белков с антигенсвязывающей активностью.