а) Значительное влияние генной мутации на болезни человека и ее связь с пародонтитом. Системные заболевания, такие как пародонтит, обусловлены сложной взаимосвязью многих генетических и негенетических факторов. В отличие от этого, моногенные заболевания, например хорея Хантингтона и муковисцидоз, полностью наследуемые. У людей, несущих причинный аллель в одном конкретном гене, неизбежно разовьется заболевание.
Синдром Папийона-Лефевра относительно уникален в группе монотонных заболеваний в том, что весомым компонентом фенотипа и определяющим клиническим признаком заболевания бывает тяжелый AgP (Toomes et al., 1999). Страдают и временные, и постоянные зубы, в результате чего пародонтит и преждевременная утрата зубов происходят в препубертатном возрасте. Кроме того, в течение первых 3 лет жизни, как правило, развивается ладонно-подошвенный кератоз, варьируя от умеренного шелушения кожи, напоминающего псориаз, до выраженного гиперкератоза. Кератоз также появляется на других участках, например локтях и коленях.
У большинства пациентов с синдромом Папийона-Лефевра развиваются как пародонтит, так и гиперкератоз. Некоторые пациенты имеют только одно из этих проявлений, причем поздний дебют или легкое течение пародонтита отмечают редко.
Причинные мутации при синдроме Папийона-Лефевра выявляют в CTSC-гене (гене катепсина С) на хромосоме 11. В настоящее время идентифицировано более 50 мутаций в этом гене. Считают, что белок, кодируемый этим геном, — катепсин С, или лизосомальная цистеинпротеаза, — служит центральным координатором активации различных сериновых протеиназ. Он экспрессируется на высоком уровне в ПЯН, альвеолярных макрофагах и их предшественниках (Rao et al., 1997).
Есть предположение, что минимальная активность катепсина С (13%) необходима, чтобы предотвратить клинические проявления синдрома Папийона-Лефевра, но точный механизм, с помощью которого измененные функции катепсина С играют роль в патогенезе препубертатного пародонтита, ассоциированного с синдромом Папийона-Лефевра, неизвестен (Hewitt et al., 2004). Возможно, катепсин С необходим для активации многих сериновых протеиназ в иммунных воспалительных клетках, в том числе катепсина G, нейтрофильных сериновых протеаз, протеиназы 3 и эластазы (Dalgic et al., 2011). Неактивные формы этих нейтрофильных сериновых протеаз приводят к нарушению регуляции иммунного ответа хозяина. Повышенная восприимчивость к инфекциям обусловлена ослаблением фагоцитарной активности нейтрофилов и нарушением функций Т- и В-клеток (Ryu et al., 2005).
Ослабленный локализованный ПЯН-ответ в воспаленных тканях пародонта приводит к AgP, скорее всего, из-за нарушенного фагоцитоза и лизиса грамотрицательных пародонтальных патогенов. Кроме того, мутация в гене CTSC, по-видимому, приводит к неспособности ПЯН уничтожать A. actinomycetemcomitans в анаэробной среде (de Haar et al., 2006).
б) Выявление генетических факторов риска развития пародонтита. Каковы общие генетические факторы риска развития пародонтита? Интересно отметить, что, несмотря на огромные усилия в течение последнего десятилетия в исследованиях генетических ассоциаций, причинные полиморфизмы генов, ответственных за пародонтит, и их патофизиологические механизмы в основном остались спорными. В этом разделе мы опишем достоинства и недостатки последних исследований, посвященных идентификации генетических факторов риска, которые лежат в основе развития пародонтита. Кроме того, обобщим и обсудим текущее состояние генетических исследований пародонтита. Начнем с того, что кратко рассмотрим результаты исследований других системных заболеваний человека.
Аналогично пародонтиту в отношении других системных заболеваний также было предложено много генетических факторов риска, и они также были спорными (Morgan et al., 2007). Эти гены рассматривали в качестве потенциальных факторов риска, но лишь немногие из них (если таковые существуют) были установлены в качестве таковых окончательно (Casas et al., 2006; Morgan et al., 2007). Некоторые факты вызывают сомнение в достоверности опубликованных отчетов, в том числе вследствие слишком малых размеров выборки, множественных сравнений подгрупп и систематических ошибок.
По-видимому, систематическая ошибка, связанная с предпочтительной публикацией положительных результатов исследования, — решающий фактор стратификации продвижения исследований в краткосрочной перспективе. Это обусовлено тем простым фактом, что намного легче опубликовать положительные результаты, чем отрицательные, что приводит к смещению публикаций в сторону накопления ложноположительных результатов в научной литературе и недостаточной представленности истинных отрицательных результатов. Тем не менее с течением времени, как правило, удается добиться истинных результатов. Эти недостатки привели к значительным сомнениям по поводу совокупной выборки предполагаемых генетических факторов риска для большинства системных заболеваний.
В 2007 г. произошел важный поворотный момент в анализе генетических факторов системных заболеваний. Эпоха исследования полногеномных ассоциаций (ИПГА; вставка 2) открылась знаковой публикацией благотворительного фонда «Wellcome Trust» (Wellcome Trust Case Control Consortium — WTCCC, 2007). Для открытия этой эпохи были необходимы две парадигмы. Во-первых, стало ясно, что опубликованные гипотезы о генах-кандидатах, как правило, не отражают ситуацию в природе, в то время как технические достижения позволяют открыть свободный от гипотез подход одновременного тестирования от 500 000 до нескольких миллионов полиморфизмов, разбросанных по всему геному, в ИПГА.
Во-вторых, было признано, что большие популяционные группы для исследований типа случай-контроль — необходимое условие для преодоления присущей гетерогенности населения, а также что потенциальные генетические факторы риска должны быть воспроизведены в больших, хорошо охарактеризованных популяциях пациентов для исключения случайных наблюдений (см. вставку 2) (Ioannidis et al., 2001).
Истинно положительные, так же как и истинно отрицательные ассоциации, можно надежно идентифицировать, только если достаточна статистическая мощность исследования клинически анализируемого населения. Статистическая значимость, в числе прочих факторов, существенно зависит от размера изучаемой популяции (см. вставку 2). Таким образом, необходимы тысячи четко определенных случаев и многие другие элементы контроля для обнаружения аллеля с небольшим эффектом, который обычно выявляют при системных заболеваниях. Реализация такой концепции в конечном итоге привела к образованию обширных международных консорциумов для набора соответствующих подходящих историй болезни и контрольных данных.
Вставка 2. Исследования генетических ассоциаций. При исследованиях с целью локализации хромосомных регионов (локусов), которые вносят вклад в восприимчивость к болезни, анализируют частоту аллельных вариантов в популяции и испытывают их совпадение с заболеванием — по сравнению с популяцией, не имеющей заболевания (контрольной группой). Цель таких исследований генетической ассоциации (или картирования ассоциации) заключается в определении того, входит ли человек, несущий одну или две копии варианта с высоким риском, в группу повышенного риска развития заболевания.
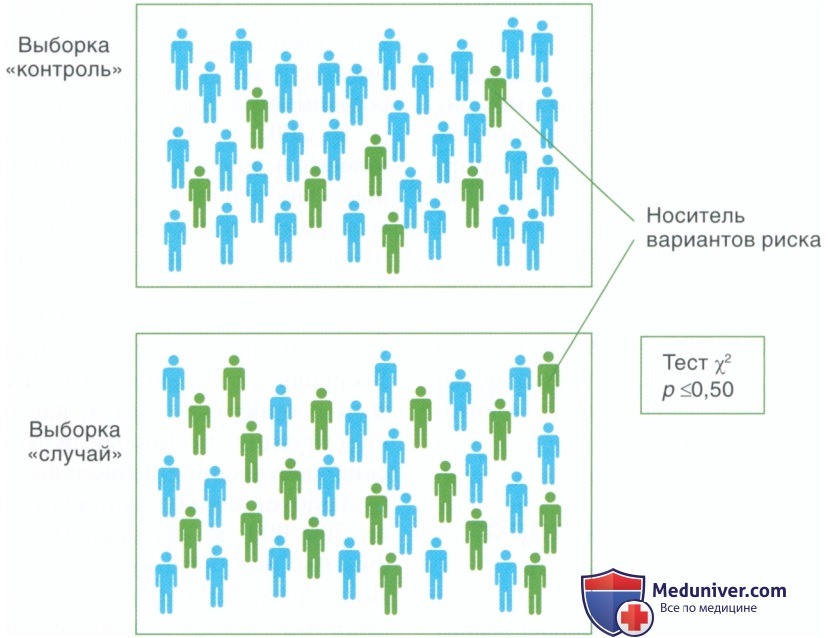
Рисунок 5. В исследованиях по типу случай-контроль сравнивают частоту аллелей: одиночного нуклеотидного полиморфизма (SNP) в двух четко определенных группах не связанных друг с другом людей: из группы контроля, не подверженных воздействию или выбранных случайным образом, и из группы «случай» с диагностированным изучаемым заболеванием. Увеличение частоты аллеля SNP или генотипа в группе «случай» по сравнению с контрольной группой указывает на тот факт, что наличие аллеля SNP может увеличить риск заболевания. Потенциальная ассоциация — лишь статистическая ассоциация, поэтому всегда требует повтора в независимой выборке. Значимость можно оценивать различными методами, но чаще всего λ2-статистику используют при анализе таблиц сопряженности, которые обеспечивают оценку отклонения от равных частот аллелей SNP в группах «случай» и «контроль» (p-значение). Исследования ассоциации также можно использовать для оценки риска развития заболевания, закрепленным за аллелем SNP, который выражается отношением шансов (ОШ). ОШ — отношение носителей аллеля к «неносителям» в группе «случай» по сравнению с группой «контроль», что дает увеличение риска заболевания для носителей по сравнению с «неносителями» (Lewis, 2002)
Принцип широко используемого исследования ассоциации по типу случай-контроль представлен на рис. 5. Данное исследование представляет собой мощный метод обнаружения ассоциации определенных аллелей с фенотипом заболевания, и его часто используют для выявления генетических факторов риска развития пародонтита.
Важным условием исследований по типу случай-контроль служит обеспечение хорошего соответствия между генетическим фоном и контролем таким образом, чтобы любое генетическое различие между ними было связано с изучаемым заболеванием, и не с необъективной выборкой. Таким образом, группы «случай» и «контроль» должны иметь аналогичное этническое происхождение. Еще одно условие — стратегия выбора группы «случай», которую разрабатывают для накопления конкретных предрасполагающих к заболеванию аллелей. Это включает усилия по минимизации фенотипической разнородности за счет строгих критериев диагностики и заставляет сосредоточиться на крайних случаях, например ограниченных очень ранним возрастом начала заболевания и/или тяжестью заболевания.
В большинстве случаев, в частности, когда общий размер выборки имеет финансовые или практические ограничения, исследования должны быть направлены на случаи наиболее тяжелых фенотипов, что может повысить статистическую значимость исследования в связи с увеличением частоты генотипа, связанного с риском (McCarthy et al., 2008). По этой же причине для определения истинного фактора генетического риска необходим анализ достаточно большой популяции при исследованиях типа случай-контроль — чтобы обеспечить необходимую статистическую достоверность в отношении первоначально исследуемой связи с заболеванием и обязательного последующего повторения наблюдений. Статистическая значимость возрастает с увеличением размера выборки и коррелирует с частотой аллеля и генетического эффекта при соответствующем варианте (Kathiresan et al., 2004).
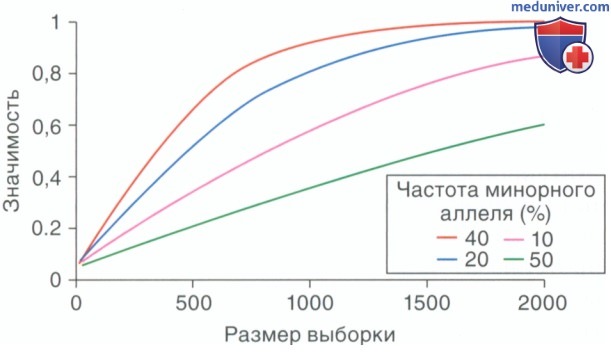
Общие варианты или варианты с высоким ОШ с большей вероятностью будут обнаружены при ИПГА, чем при редких вариантах или не очень выраженных проявлениях патологии (рис. 6).
Рисунок 6. Статистическая значимость в зависимости от размера выборки, частоты аллеля и отношения шансов (ОШ). К примеру, чтобы определить генетический вариант риска с частотой минорного аллеля 20% в общей популяции, для достижения необходимой статистической значимости 0,8 необходимы = 1000 больных и 2000 представителей контрольной группы. [Статистическая значимость была рассчитана, как описано Dupont и Plummer (1998), для среднего ОШ 1,3, и были рассмотрены в 2 раза больше людей из групп как «контроль», так и «случай». Коэффициент 0,8 считали статистически значимым.]
Тем не менее большинство связанных с заболеванием генетических вариантов незначительно повышает восприимчивость к нему, поэтому при не очень ярких проявлениях болезни необходимо >1000 хорошо выраженных «случаев» и, по крайней мере, такое же количество «контроля» — для достижения достаточной статистической значимости в отношении генетического эффекта.
Результаты исследований по типу случай-контроль следует считать лишь статистическими сопоставимыми, описывающими различия частот аллелей в двух независимых выборках. Их не нужно рассматривать в качестве причинных ассоциаций. Предварительный порог достоверности 0,05 означает, что 1 из каждых 20 тестируемых аллельных вариантов будет соответствовать обычно задаваемому значению р<0,05. Частоты аллелей в независимых друг от друга выбранных популяциях также подвержены случайным колебаниям (случайные переносы аллеля по горизонтали и между популяциями, не зависящие от отбора).
По этим причинам повторяемость первоначальных результатов в последующих исследованиях — золотой стандарт для генетических исследований ассоциации. Следует отметить, что повторяемость необходима в отдельной выборке случай-контроль того же фенотипа (критерий диагностики) и той же этнической принадлежности.
Повторение исследования с выборками из различных этнических групп, различными диагностическими критериями или независимыми случаями, но с тем же контролем, нельзя рассматривать как повторяемость, поскольку такие ситуации не соответствуют критериям первоначального обнаружения связи. Только подтверждение четкого повтора свидетельствует о полезности исследования для дальнейшей проверки первоначально выявленных ассоциаций при различных субфенотипах или в различных этнических группах.
Поскольку гены обычно представляют собой мозаики различных гаплотипов (в основном от бедных до умеренно неравновесных по сцеплению), информация о значимости одного гаплотипа практически не дает данных о роли других гаплотипов в пределах этого гена (Slatkin, 2008). Таким образом, нужна полная информация о гаплотипах интересующего гена, чтобы сделать вывод о наличии или отсутствии связи этого гена с изучаемым явлением (Slatkin, 2008).
1. Исследование ассоциаций генов-кандидатов. До середины прошлого десятилетия исследования отобранных генов-кандидатов на основе обзоров литературы и предполагаемых патофизиологических путей были самой важной стратегией для идентификации генов риска, которые способствуют развитию заболевания. Основной недостаток исследований генов-кандидатов — требование априорной гипотезы о причастности гена к риску заболевания и наличии функционального варианта в рамках этого конкретного гена (Wilkening et al., 2009). По существу, есть две разные стратегии отбора генов-кандидатов, которые зависят от поставленного вопроса. Когда возникает вопрос, есть или нет специфический локус в пределах регуляторного сигнального пути, который участвует в повышении генетического риска пародонтита, либо есть функциональное свидетельство о влиянии варианта гена на другие заболевания, разумно выбрать гены из этих путей или специфических вариантов.
Такой подход позволит определить, действительно ли выбранные гены несут генетические варианты, повышающие риск заболевания.
Другой вопрос, который обращается к классической задаче молекулярной генетики, сложнее: какие конкретные гены и пути влияют на риск развития заболевания? Поскольку формулировка гипотезы о выборе гена-кандидата полностью зависит от текущего знания молекулярных биологических механизмов заболевания, сотни локусов и/или генов, которые могут иметь влияние на болезнь, не будут выбраны, потому что их функции неизвестны или имеют отношение к пути, который еще не связывали с заболеванием. В свете того, что знания об этих генах неполные, выбор генов-кандидатов произволен. Соответственно, большинство связей, заявленных в этих научных отчетах, невозможно повторно подтвердить в других исследованиях. Очевидно, что это не исключает нахождения истинной положительной ассоциации с заболеванием, если правильный ген-кандидат был выбран заранее, но при таком подходе невозможно идентифицировать неизвестные ранее гены, которые приводят к болезни.
2. Исследования полногеномных ассоциаций. В противоположность изложенному выше, ИПГА обеспечивают объективный и свободный от гипотез подход. Большое количество полиморфизмов (в настоящее время от 500 000 до >1 000 000 маркеров), распределенных по всему геному, служат в качестве прокси-объектов для множества других полиморфизмов при неравновесии по сцеплению. Тем не менее тестирование полиморфизмов по всему геному также влечет за собой проблемы. Во-первых, если один из каждых 20 маркеров, тестируемых по одному случайным образом, дает значение р<0,05, вероятность статистических ошибок возрастает с увеличением количества одиночных испытаний ассоциации SNP — так называемые ошибки 1-го типа (ложноположительные результаты ассоциации).
Если независимо проверено 500 000 маркеров или более, значение р, полученное из λ2-статистики, должно быть скорректировано для многократного тестирования. Это решается путем установки полногеномного порога значимости путем коррекции количества проведенных тестов (Balding, 2006). Текущий стандарт для признания статистической значимости на общегеномном уровне представляет собой комбинированное значение р (в том числе «первоначальное открытие» ИПГА и репликация когорт) <5х10-8 (Manolio, 2010). Тем не менее размеры выборок, которые необходимы для достижения таких порогов значимости, могут быть нереалистичными для изучения менее распространенных заболеваний. В результате недостаток статистической мощности — основной фактор, который приводит к ошибкам 1-го и 2-го типов (ложноположительные и ложноотрицательные результаты), т.е. неспособности обнаружить истинную ассоциацию.
Первую волну ИПГА в течение 2007-2010 гг. часто называют генетической золотой лихорадкой в генетике человека, когда были выявлены многие из общих генетических факторов риска системных заболеваний человека (т.е. генетические варианты с >5% преобладанием в данной популяции). Несмотря на то обстоятельство, что эти исследования оказались весьма успешными в определении наиболее распространенных генетических вариантов восприимчивости, они могли объяснить лишь часть основной генетической наследуемости (Maher, 2008; Manolio et al., 2009). Оказалось, что возможности отдельного исследования, охватывающего >1000 случаев, по-прежнему ограничены в плане обнаружения небольших или незначительных эффектов, связанных с разными вариантами заболевания.
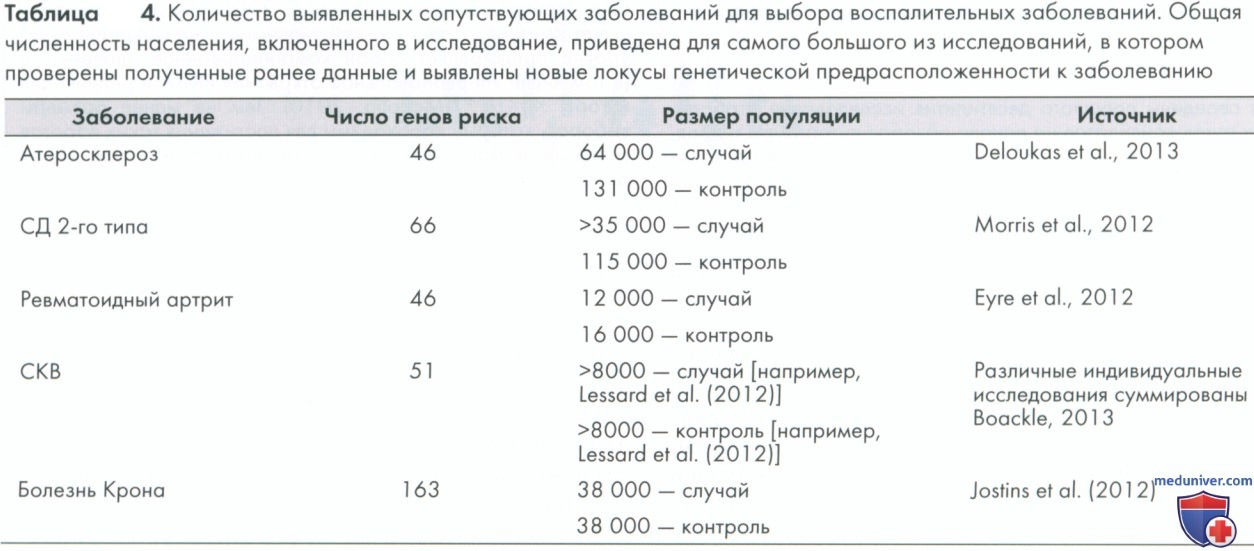
Для того чтобы обнаружить такие варианты с умеренным эффектом, а также редкие варианты с большим эффектом на индивидуальном уровне, необходимо гораздо больше анализируемого населения. С этой целью исследователи первого ИПГА объединили свои усилия и комбинировали данные для метаанализа, который в конечном счете включал более десятка тысяч случаев контроля. Таким образом, было открыто большинство общих генетических факторов риска, сопутствующих развитию заболеваний пародонта, таких как СД 2-го типа, ишемическая болезнь сердца или ревматоидный артрит. В табл. 4 предоставлен краткий обзор результатов последних лет с точки зрения количества выявленных локусов генетического риска основных системных воспалительных заболеваний, некоторые из которых связаны с сопутствующими заболеваниями при пародонтите, и количества случаев и контроля, используемых в этих исследования. Хотя выявлено много локусов риска для различных заболеваний, ожидают, что их количество увеличится в ближайшие годы.
Интересно, что большинство из выявленных генов не рассматривали до этого как вероятные гены-кандидаты. Эти данные подтверждают полигонный характер системных заболеваний, а также небольшой патогенетический вклад SNP в отдельные гены. Мы предполагаем, что это справедливо и для пародонтита.
Большинство генетических исследований ассоциаций, которые были разработаны для выяснения генетических факторов риска развития пародонтита, сосредоточено на различных генах-кандидатах, отобранных в соответствии с их ролью в иммунной системе (например, гены IL и семейств толл-подобных рецепторов), деструктивных процессах в тканях (например, матричная металлопротеиназа) или различных механизмах обмена веществ (Loos et al., 2005; Laine et al., 2010, 2012). По причинам, аналогичным описанным для других системных заболеваний в пре-ИПГА эпоху, несмотря на многочисленные усилия в области генетических исследований ассоциаций, генетические факторы риска развития пародонтита и их патофизиологические эффекты во многом остаются неоднозначными.
Неоспоримы лишь немногие гены. Высказаны значительные сомнения по поводу совокупной выборки предполагаемых генетических факторов риска. Чтобы уточнить роль комплексного набора генов, которые были в центре внимания недавнего генетического исследования по пародонтиту, провели крупномасштабное, воспроизводимое и детальное исследование 23 генов (AB0, CCR5, FCGR2A, FCGR2C, FCGR3A, FCGR2B, FCGR3B, IL-1b, IL-2, IL-6, IL-10LTA, ММР-9, NOD2, TLR-2, TLR-4, РДР, CD14, IL-1A, IL-1RN, TNFRSF11B, IFNGR1, L-селектпина) у 600 пациентов из Германии с AgP и 1440 человек контрольной группы также из Германии со средним охватом SNP из <5000 пар оснований (Schafer et al., 2013). За исключением IL-10, это исследование не выявило сильной связи между тестируемыми генами и их регуляторными областями с AgP, и авторы предположили, что позитивные ассоциации с пародонтитом, которые ранее были зарегистрированы для этих генов, скорее всего, вызваны ошибками 1-го типа.
Такая бездоказательность роли классических генов-кандидатов в развитии пародонтита — подчеркивают авторы далее — требует осторожности в интерпретации генетических ассоциаций и существенной проверки сообщений о генетических факторах риска.
Некоторые гены все еще можно считать истинными генетическими факторами восприимчивости к пародонтиту, если доказательства этого многократно подтверждаются в повторных исследованиях по типу случай-контроль независимых, достаточно больших групп населения.
в) ANRIL, CAMTA1/VAMP3, GLT6D1, СОХ-2 и NPY (вставка 3):
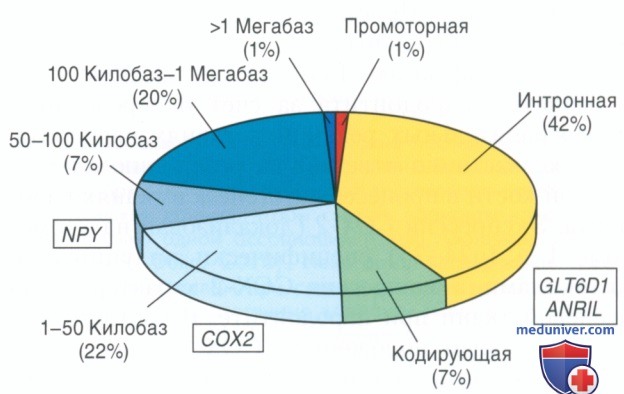
Вставка 3. Геномное расположение генетических вариантов риска. Недавнее обсуждение в генетических обзорах системных заболеваний, в том числе пародонтита, позволяет предположить, что общие генетические варианты риска, вероятно, следует искать не в кодирующих областях классических генов-кандидатов, а внутри регуляторных элементов непредвиденных генов и хромосомных областей. Рис. 7 демонстрирует, что большинство распространенных геномных вариантов человека, которые тесно связаны более чем с 400 общими заболеваниями и признаками, находятся в пределах регуляторных активных областей. Это также верно для расположения генетических вариантов риска развития пародонтита, которые являются интронами для обоих ANRIL и GLT6D1, лежащими несколькими тысячами пар нуклеотидов вверх и вниз от белок-кодирующих областей СОХ-2 и NPY.
Рисунок 7. Хромосомное расположение связанных с заболеванием одиночных нуклеотидных полиморфизмов в общегеномных исследованиях ассоциаций различных системных заболеваний. В среднем 64% ассоциированных с заболеваниями вариантов расположено в пределах интронов на 1-50 килобаз вверх или вниз по протяженности гена. Расположение проверенных паро-донтит-ассоциированных вариантов в соответствии с этим наблюдением (изображено оранжевым).
Как правило, количество вариантов, связанных с болезнью и характерными особенностями, значительно увеличивается в пределах участков, физиологически или патогенетически соответствующих регуляторным областям. Интересно, что они также группируются в пределах известных регуляторных путей и представляют функциональные сети, которые концентрируются на конкретных транскрипционных регуляторах. Результаты последних ИПГА указывают на широко распространенное вовлечение регуляторных вариаций ДНК в общие заболевания человека, а также на импликацию многочисленных транскрипционных регуляторов в восприимчивости к болезни и патогенезу.
1. ANRIL. В настоящее время наиболее воспроизводимые доказательства риска развития пародонтита отмечены для гена ANRIL [«бессмысленная некодирующая РНК в локусе INK4» (Pasmant et al. 2007); образец в GenBank №DQ485453; код CDKN2B-AS1 (CDKN2B бессмысленная РНК 1) на хромосоме 9р21.3], как показано в трех независимых северо-западных европейских популяциях (Schafer et al., 2009; Ernst et al., 2010) и одной турецкой популяции (Schafer et al., 2013) при изучении AgP, а также в одной популяции при изучении хронического пародонтита (Schafer et al., 2011).
Интересно, что ANRIL также считают наиболее важным генетическим фактором риска развития ИМ, как это определено в различных ранних ИНГА по ишемической болезни сердца (ИБС) (McPherson et al., 2007; Samani et al., 2007; WTCCC, 2007). Этот ген кодирует десенсибилизирующую кодирующую большую небелковую молекулу РНК, А-126,6 кб полная длина транскрипта 19 дифференциально сплайсированных экзонов, с образованием различных молекул РНК различной длины.
Несмотря на обширные исследования в течение последних нескольких лет, молекулярная функция ANRIL остается очень мало изученной, и природа причинных вариантов до сих пор четко не определена. Пока неизвестно, как генетическая предрасположенность к различным заболеваниям, связанным с этим локусом, отражается на физиологических функциях. Пришли к согласию, что ANRIL оказывает отрицательное регулирующее воздействие на экспрессию ингибиторов соседней циклин-зависимой киназы — генов CDKN2A и CDKN2B (Visel et al., 2010; Yap et al., 2010).
2. CAMTA1/VAMP3. В соответствии с наблюдениями, длинные некодирующие РНК часто играют роль в трансрегуляторной экспрессии генов (Pandey et al., 2008; Mercer et al., 2009). Было показано, что снижение экспрессии ANRIL коррелирует со снижением уровня экспрессии отдаленных генов ADIPOR1 (рецептор адипонектина 1-го типа; хромосома 1), VAMP3 (везикуло-ассоциированный мембранный белок 3; хромосома 1) и C11ORF10 (хромосома И, открытая рамка считывания 10; хромосома И) (Bochenek et al., 2013). Белок VAMP3 принадлежит к семейству VAMP (семейству синаптобревина) и играет роль в фагоцитозе, где VAMP3 опосредует, например, доставку TNFa к поверхности клетки (Murray et al., 2005). Расположенный в 2 килобазах перед VAMP3, это чрезвычайно большой ген САМТА1 (кальмодулин-связывающий активатор транскрипции 1), охватывающий >1 мегабаз.
В ИПГА по колонизации пародонтальных патогенов сообщают о большом участке САМТА1/ VAMP3-области, тесно связанной с увеличением количества патогенных бактерий в полости рта (Divaris et al., 2012). Поиски потенциальных паро-донтит-ассоциированных вариантов в пределах САМТА1/VAMP3 определили несколько SNP, расположенных в этой патоген-ассоциированной области, явно связанных с AgP (Bochenek et al., 2013). Интересно, что эта область значительно увеличивает риск ИБС — наблюдение, которое впоследствии было подтверждено в метаанализе 13 ИПГА ИБС, содержащем 21 033 пациента с ИБС и 44 065 людей контрольной группы европейского происхождения (Bochenek et al., 2013). Следовательно, CAMTA1/VAMP3-регион — вторая подтвержденная область общего локуса генетического риска пародонтита и ИБС.
3. GLT6D1. Другой ген, ассоциированный с риском развития AgP, GLT6D1, также картированный на хромосоме 9, в локусе 9q34.3, был идентифицирован в первом ИПГА по пародонтиту (Schafer et al., 2010b). Он кодирует неизвестный белок, относящийся к семейству белков, который характеризуется гликозилтрансферазой домена-1. Установлено, что GLT6D1 преимущественно экспрессируется в десне и Т-клетках. Молекулярная функция этого гена до сих пор неизвестна, но секвенирование и последующая молекулярная биологическая характеристика основных связанных с ним генетических полиморфизмов предполагают нарушенный сайт связывания фактора транскрипции GATA3 как вариант, вызывающий повышение риска заболевания.
4. СОХ-2. Метаболический белок СОХ-2 преобразует арахидоновую кислоту в простагландин Н2, предшественник PGE2. Простагландины — группа ключевых воспалительных медиаторов иммунного ответа на инфекцию. PGE2 играет важную роль в развитии пародонтита за счет опосредования провоспалительных реакций в тканях пародонта, а также частично отвечает за резорбцию альвеолярной кости в процессе патогенеза в тканях пародонта. Экспрессия СОХ-2 (локализован на хромосоме 1 в 1q24—25) специфически индуцируется цитокинами, а экспрессия СОХ-2 характерна для десневой ткани при пародонтите. В соответствии с центральным положением СОХ-2 в регуляции уровней простагландина Н2 и специфичной активации цитокинов ген СОХ-2 подвергался разнообразным кандидат-SNP ассоциативным исследованиям при различных системных заболеваниях, и полиморфизмы в гене СОХ-2 были связаны с повышенной восприимчивостью к различным воспалительным заболеваниям.
При исследованиях пародонтита дизайна случай-контроль в тайваньских и китайских популяциях было независимо показано, что та же генетическая область связана с тяжелой формой пародонтита (Но et al., 2008; Xie et al., 2009), и эта ассоциация была впоследствии подтверждена в европейской популяции с AgP (Schafer et al., 2010a).
Таким образом, в этих исследованиях представлены убедительные доказательства того, что ограниченная хромосомная область в направлении к СОХ-2, возможно, несет один или несколько вариантов риска развития тяжелого пародонтита в популяциях различного этнического происхождения.
5. NPY. Нейропептид Y (NPY) оказывает иммуномодулирующее действие, которое, как полагают, смещает баланс между провоспалительными Т-хелперами типа 1 (ТЫ) и противовоспалительными Т-хелперами типа 2 (Th2) в сторону последних. Связывание NPY с рецепторами Y1 в различных иммунных клетках предположительно отвечает за активацию противовоспалительного ответа Th2 (Bedoui et al., 2003). Следовательно, NPY потенциально важен для координации воспаления и метаболизма костной ткани, которые играют центральную роль в патогенезе пародонтита (Lundy et al., 2009). Соответственно наличие рецепторов NPY типа Y1 верифицировано в десневой ткани человека, а самого NPY — в GCF, причем со значительно более высоким уровнем NPY в GCF здорового пародонта по сравнению с участками пародонтита (Lundy et al., 2009).
В первом ИПГА обнаружена ассоциация с тяжелым хроническим пародонтитом нижней кодирующей области NPY (хромосома 7) в большой выборке европейско-американских представителей (Divaris et al., 2013). Второе ИПГА показало, что при систематическом анализе генно-половых взаимодействий в случаях AgP и в контрольных группах в Германии роль полового диморфизма генетических вариантов NPY была связана с AgP (Freitag-Wolf et al., 2014). Интересно отметить, что зависимые от иола эффекты NPY были ранее описаны у мышей. Мыши с утраченной функцией NPY продемонстрировали различные анксиогенные ответы в поведенческих тестах у самцов и самок, что свидетельствует о роли NPY в половом диморфизме поведенческих реакций на стресс.
Кроме того, желудочно-кишечное воспаление, как известно, увеличивает тревогу, выражаемую секс-зависимым образом, что вызывает различные поведенческие реакции на стресс женских и мужских особей мышей, дефектных по NPY (Painsipp et ah, 2011). NPY активирует гипоталамо-гипофизарно-надпочечниковую систему и модулирует висцеральные ответные реакции на стресс, опосредованные через пути кортикотропин-рилизинг-гормона (Dimitrov et al., 2007). Кроме того, NPY — сильный анксиолитик (Karl et al., 2008), действующий через Y1-рецепторы NPY в миндалине, ингибируя передачу сигналов кортикотропин-рилизинг-гормона и прерывая поведенческий стресс и тревожный ответ.
Гены ANRIL, GLT6D1, СОХ-2 и NPY - неоспоримые гены риска развития пародонтита (ANRIL, СОХ-2 и NPY — для AgP и хронического пародонтита, GLT6D1 — только для AgP), что подтверждается повторением данных в независимых больших группах населения. Очевидно, что эти данные не объясняют полностью наследственность пародонтита, и еще предстоит открыть множество других генов, генетических элементов, а также их вариантов (вставки 4, 5).
Вставка 4. Текущие и будущие стратегии для выявления полного спектра генетических изменений при пародонтите. Основная парадигма, используемая в недавно завершенном этапе ИПГА в генетическом распределении часто встречаемых воспалительных заболеваний, — классифицировать общие факторы риска. Эти ИПГА успешно представлены вариантами с частотой, значительно превышающей 5% общей популяции. Для выявления менее распространенных вариантов целесообразно расширение ИПГА с большим охватом редких вариантов. Это направление мысли подчеркнул международный консорциум «Проект 1000 геномов» (www.1000genomes.org), который расширил каталог известных человеческих вариантов с частотой около 1%. Новые ИПГА будут вносить в каталог менее распространенные варианты с частотами аллелей 1-5% и выявлять некоторые новые ассоциации редких вариантов.
Тем не менее, чтобы захватить редкие варианты с частотами аллелей <1%, целые геномы или целые экзомы должны быть секвенированы, чтобы обеспечить достаточный охват для чтения всех вариаций ДНК, в том числе общих и редких вариантов, включенных в монотонные и системные заболевания. В конце концов исследования роли наследственной изменчивости в развитии заболеваний будут включать секвенирование целого генома всех испытуемых. Такие исследования будут осуществляться подобно ИПГА, с очень большими размерами выборки, которые обеспечат достаточное количество статистических данных, чтобы связать варианты на основе данных только одной ассоциации (Cirulli, Goldstein, 2010; Janssens, van Duijn, 2010). Некоторые исследования уже показали, что в целом при секвенировании экзома можно идентифицировать патогенные варианты (Choi et al., 2009; Ng et al., 2010a, b). Тем не менее пока полное геномное секвенирование не станет достаточно дешевым, чтобы использоваться в больших выборках, исследователи будут применять различные другие модели для выявления общих и редких этиологических вариантов.
1. Отбор семей, в которых присутствуют несколько заболевших (секвенирование на базе семьи), и отбор людей, которые находятся в крайних положениях распределения признака (экстремальная черта моделей).
2. Секвенирование полного экзома или полноэкзомное генотипирование массивов, которые составляют комплект почти полного спектра известных вариантов кодирования в человеческом экзоме (Cirulli, Goldstein, 2010).
Вставка 5. Перспективы. Недавние ИПГА предоставили ценную информацию о генетической основе системных заболеваний и многих выявленных генах, обусловливающих восприимчивость к болезни. В 2014 г. в каталог ИПГА Национального института генома человека США (NHGRI; www.genome.gov/gwastudies) были включены 14 769 SNP с геномными ассоциациями с >600 признаками (Welter et al., 2014). Тем не менее эти варианты объясняют лишь небольшую часть наследуемости изучаемых признаков (Frazer et al., 2009). Это произошло отчасти из-за статистических ограничений, которые присущи ИПГА и вообще допускают только обнаружение общих вариантов риска. В настоящее время идет обсуждение ситуаций, при которых наследуемость склонности к заболеванию практически очевидна, но не подтверждена в исследованиях: какие методы исследования помогут выявить неизвестные генетические факторы риска (Eichler et al., 2010).
Несколько лет назад, когда началось проведение ИПГА, было широко распространено мнение, что системное заболевание чаще связано с небольшим количеством общих вариантов, каждый из которых обусловливает несколько процентов риска в популяции: это называют общим заболеванием — общим вариантом гипотезы (Pritchard, Сох, 2002). Данная модель бросает вызов так называемой «проблеме скрытой наследуемости», которая описывает наблюдение, что локусы, обнаруженные при ИПГА, почти без исключения вносят лишь ограниченный вклад в заболевание (Maher, 2008). В этом контексте Gibson (2011) заявил, что «это было просто не так, как бывает, когда несколько десятков локусов, оказывающих умеренный эффект, и промежуточная частота каждого обусловливают несколько процентов риска заболевания в популяции — как это обычно регистрировали при скрещивании или в генеалогии». Таким образом, генетическая дисперсия была отнесена преимущественно к одной из двух следующих причинных моделей болезни.
1. Большое количество малоэффективных распространенных вариантов по всему спектру частот аллеля вызывает генетическую восприимчивость к заболеванию. В основе этой гипотезы лежит предположение, что общие генетические варианты — один из основных источников генетической дисперсии или восприимчивости к болезни, а сотни или даже тысячи различных локусов вносят свой вклад в каждый конкретный случай. Локусы, обнаруженные при ИПГА, — лишь наиболее эффективные и масштабные в распределении по Пуассону или другому подобному распределению (Gibson, 2011). Если 10 общих вариантов вызывают 10% риска в популяции, остальная часть обусловлена множеством вариантов, так что каждый вызывает значительно меньше 1% риска и имеет генотип относительного риска <1,1 (Gibson, 2011). Заболевание вызвано сочетанием генетических, экологических и поведенческих факторов (Feldman, Lewontin, 1975; Eichler et al., 2010). Эту модель называют «бесконечно малой моделью» (Visscher et al., 2008).
2. В альтернативном варианте в «редких аллельных моделях» большая часть генетической дисперсии происходит из-за пенетрантных вариантов с частотами аллелей <1%, но это сильно повышает генетический риск. Множество различных редких вариантов имеет большое влияние на фенотипическую вариабельность (Cirulli, Goldstein, 2010). Каждый из этих вариантов обусловливает большую часть риска в подгруппе случаев, но не объясняет в полной мере различия в общей популяции. И все варианты невозможно обнаружить при стандартном ИПГА. Несмотря на это, тяжесть заболевания в значительной степени вызвана редкой чувствительностью генотипа, он может быть модифицирован с помощью воздействия среды (что способно привести к изменению пенетрантности генетических эффектов как редких, так и общих вариантов), эпистаза или эпигенетических факторов (Bodmer, Bonilla, 2008). Однако недавнее крупномасштабное исследование показало, что редкие варианты кодирующей области в известном локусе риска аутоиммунных заболеваний играют незначительную роль в общей аутоиммунной восприимчивости к болезни (Hunt et al., 2013).
Альтернативное исследование, в котором использовали дрожжи в качестве модели для изучения скрытой наследуемости системных признаков, показало, что взаимодействие известных общих генетических вариантов могло бы объяснить почти весь аддитивный вклад в наследственные изменения (Bloom et al., 2013). Эти результаты в настоящее время складываются не в пользу гипотезы редко встречаемой синтетической полногеномной ассоциации.
По всей вероятности, каждый из этих генетических механизмов внес свой вклад в различные заболевания, но пока недостаточно данных, чтобы прийти к одному мнению, несмотря на множество сильных аргументов в пользу или против каждой из этих двух моделей (Gibson, 2011). Дебаты о вкладе генетической изменчивости в болезнь в ближайшие годы будут сосредоточены на том, насколько влияют друг на друга общие и редкие варианты (Schork et al., 2009). Очевидные гипотезы утверждают, что общая изменчивость влияет на экспрессию и активность генов на молекулярном уровне, создавая исходную восприимчивость к заболеванию, которая затем дополнительно модифицируется редкими вариантами с существенными эффектами (рис. 9).
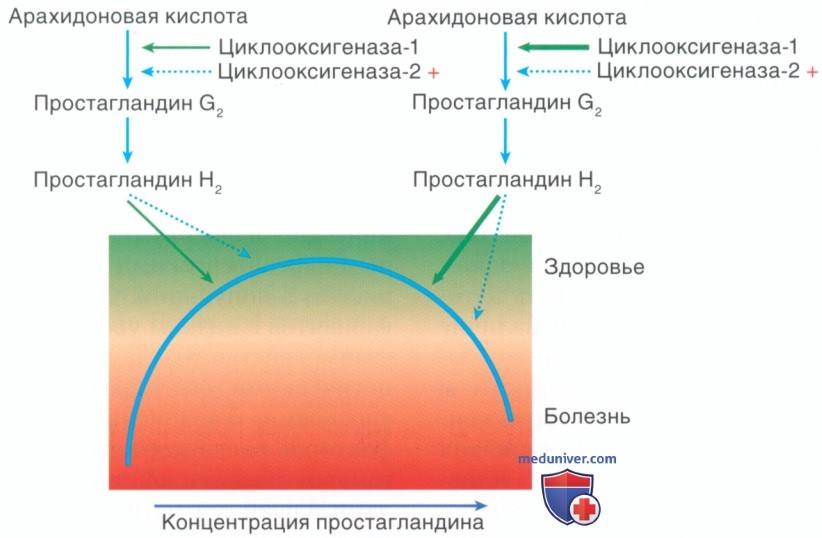
Рисунок 9. Предполагают, что общие варианты влияют на экспрессию и активность генов, создавая исходную восприимчивость к заболеванию, которая затем дополнительно модифицируется более редкими вариантами с более значительным эффектом. Простагландины вызывают каскад биохимических реакций после последовательного окисления арахидоновой кислоты посредством циклооксигеназ СОХ-1 и СОХ-2 и терминальной простагландин-синтазы. В то время как СОХ-1 отвечает за базовые уровни простагландинов, СОХ-2 производит простагландины путем специфической стимуляции по сценарию воспаления тканей пародонта. Полукруг представляет собой диапазон концентраций простагландинов при поражении у данной популяции. Концентрация простагландинов зависит от взаимодействия отдельных генетических структур и индивидуальных физиологических состояний и окружающей среды. Концентрации простагландина на нижнем и верхнем концах связаны с болезнью, в то время как промежуточная концентрация — физиологическая и совместима со здоровьем. На этой гипотетической иллюстрации генетическая изменчивость располагается на пути синтеза простагландинов у некоторых людей, имеющих более низкие уровни простагландинов (слева нормальная активность СОХ-1, обозначенная зеленой горизонтальной стрелкой от СОХ-1), чем у других (справа генетическая изменчивость СОХ-1, которая устанавливает исходную восприимчивость к болезням, указана толстой зеленой горизонтальной стрелкой от СОХ-1). Индивидуальная изменчивость исходной чувствительности все еще в пределах здорового диапазона. Воздействие дополнительного варианта, который повышает синтез СОХ-2 (обозначено знаком «+» и синими пунктирными стрелками) в результате воспалительной стимуляции, в условиях этой лабильности смещает людей с высокой концентрацией простагландинов и генетической исходной восприимчивостью (тех, что справа) в сторону развития пародонтита (в красную зону опасности). Люди с низкой концентрацией простагландинов (слева) могут приспосабливаться к генетической изменчивости и оставаться в зеленой зоне безопасности.
Рис. 9 демонстрирует, что заболевание проявляется в ответ на превышение порогового уровня раздражителя, который накладывается на непрерывную физиологическую характеристику.
Поскольку заболевание развивается только у доли генетически предрасположенных и/или патоген-подверженных людей, простых генетических объяснений индивидуальной восприимчивости к хроническим воспалительным заболеваниям, таким как пародонтит, пока нет. Задачи дальнейших исследований, помимо идентификации, будут заключаться как в выяснении максимального количества истинных факторов восприимчивости, так и в распознавании соответствующих паттернов в пределах полученных данных, другими словами, моделировании эффектов SNP-SNP-взаимодействий (Renz et al., 2011).