Знание клеточных и молекулярных механизмов развития сердца необходимо для понимания сути ВПС и еще более важно при разработке стратегий профилактики, будь то клеточная или молекулярная терапия, или кардиологические вмешательства у плода. Пороки сердца традиционно группируют по общим морфологическим признакам: напр., аномалии выносящих трактов (конотрункальные ВПС, такие как тетрада Фалло и общий артериальный ствол) и дефекты АВ-перегородки (первичный ДМПП, полная форма открытого АВ-канала). Эти морфологические категории могут изменяться или в конечном итоге вытесняться новыми категориями по мере получения новой информации о генетических основах ВПС.
а) Ранний морфогенез сердца. У раннего пресомитного эмбриона первые идентифицируемые кластеры сердечных клеток-предшественников располагаются в мезодерме передней боковой пластинки по обе стороны от центральной оси эмбриона; эти кластеры к 18-му дню в/утробного развития образуют парные сердечные трубки. Зона сердечных клеток-предшественников формируется за счет сбалансированного градиента положительных и отрицательных сигналов от тканей, окружающих сердечные мезодермальные клетки, сигналов от окружающих вентральных/латеральных тканей, способствующих кардиогенезу с помощью сигнальных молекул, таких как BMP (костный морфогенетический белок) и FGF8 (фактор роста фибробластов 8), и сигналов от дорсальных/медиальных структур, таких как участники пути Wnt/β-катенин, ингибирующие кардиогенез.
Кардиогенные сигналы активируют экспрессию кардиоспецифических факторов транскрипции (напр., Tbx, GATA, Nkx2,5), стимулирующих экспрессию сердечных генов. Парные трубки сливаются по средней линии на вентральной поверхности эмбриона, образуя примитивную сердечную трубку к 22 дням в/утробного развития. Эта прямая сердечная трубка состоит из внешнего миокардиального слоя, внутреннего эндокардиального слоя и среднего слоя внеклеточного матрикса, называемого сердечным гелем.
Выделяют две разл. клеточные линии: первое поле сердца (регулируемое в основном Nkx2,5) является источником клеток-предшественников для ЛЖ; второе поле сердца (регулируемое главным образом Isll) становится источником клеток-предшественников для предсердий и ПЖ. Премиокардиальные клетки, включая эпикардиальные клетки и клетки из нервного гребня, продолжают миграцию в область сердечной трубки. Регуляция этой ранней фазы морфогенеза сердца частично контролируется взаимодействием специфических сигнальных молекул или лигандов, обычно экспрессируемых одним типом клеток, со специфическими рецепторами, обычно экспрессируемыми др. типом клеток.
Информация о позиционировании передается развивающейся сердечной мезодерме с помощью таких факторов, как ретиноиды (изоформы витамина А), которые связываются со специфическими ядерными рецепторами и регулируют транскрипцию генов. Миграция эпителиальных клеток в развивающуюся сердечную трубку регулируется белками внеклеточного матрикса (напр., фибронектином), которые взаимодействуют с рецепторами клеточной поверхности (интегринами). Клиническое значение этих путей передачи сигналов подтверждается спектром сердечных тератогенных эффектов, вызываемых ретиноидоподобным ЛС изотретиноином.
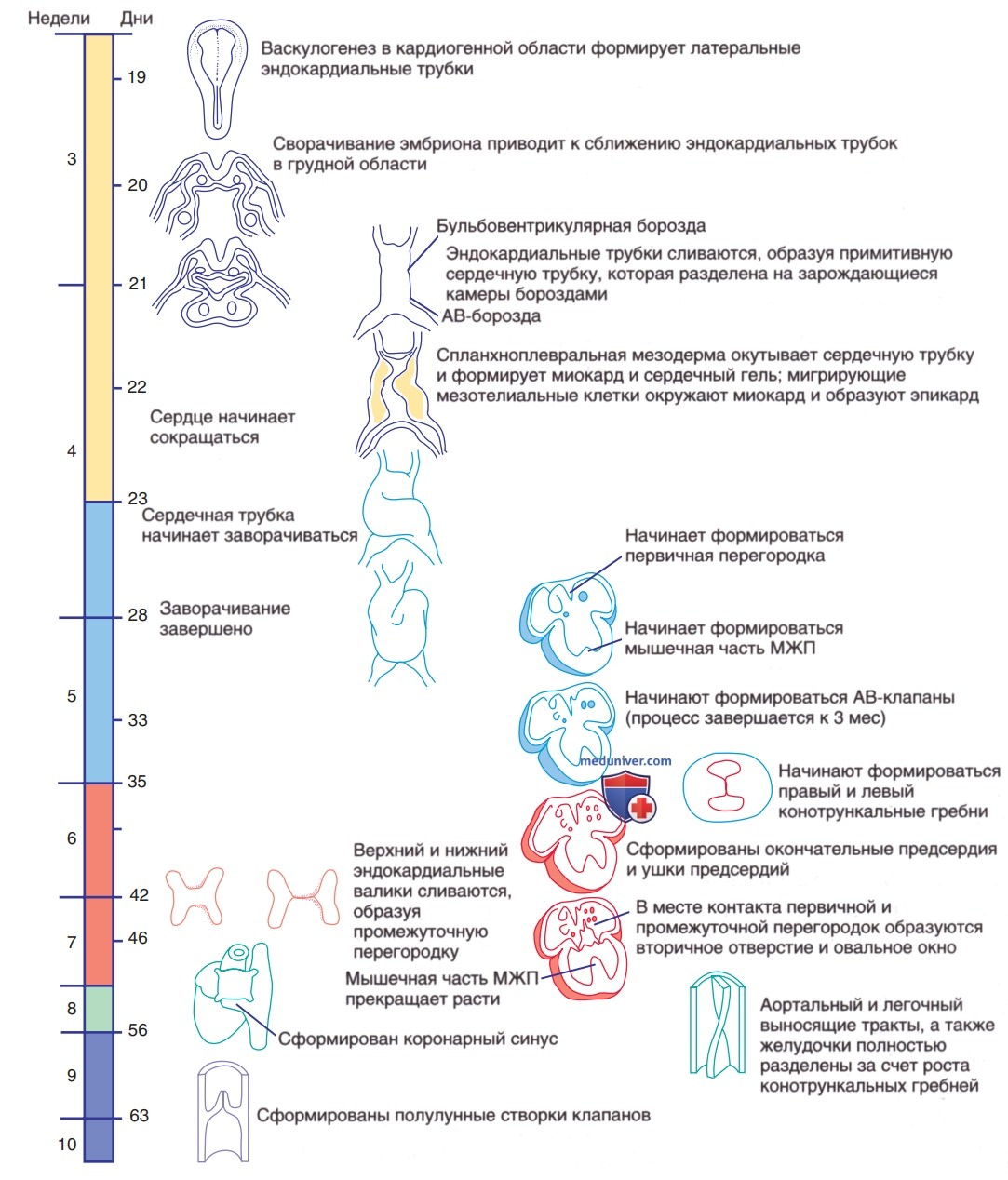
Уже на 20-22-й день в/утробного развития, до формирования сердечной петли, эмбриональное сердце начинает сокращаться с фазами сердечного цикла, которые удивительно похожи на фазы в зрелом сердце. Первоначально морфологи идентифицировали сегменты сердечной трубки, которые, как полагали, соответствовали структурам зрелого сердца (рис. 1): венозный синус и предсердие (правое и левое предсердия), первичный желудочек (ЛЖ), сердечная луковица (ПЖ) и артериальный ствол (аорта и ЛА). Однако эта модель слишком упрощенная.
Рисунок 1. Сроки морфогенеза сердца
В ранней сердечной трубке присутствуют только трабекулярные (наиболее богатые мышечными элементами) части миокарда ЛЖ; клетки, которые станут приточным трактом ЛЖ, мигрируют в сердечную трубку на более поздней стадии (после начала формирования петли). Еще позже появляются примордиальные клетки, дающие начало большим артериям (артериальный ствол; truncus arteriosus), включая клетки из нервного гребня, которые не обнаруживаются до тех пор, пока формирование сердечной петли не завершится. Факторы транскрипции для камер сердца участвуют в дифференцировке предсердий от желудочков, а также в ПЖ и ЛЖ.
Основной фактор транскрипции спираль-петля-спираль (bHLH) dHAND экспрессируется в развивающемся ПЖ; патология этого гена или др. факторов транскрипции, таких как факторы усиления транскрипции миоцитов 2С (MEF2C) у мышей, приводит к гипоплазии ПЖ. Др. генетические маркеры клеток второго поля сердца (ранний ПЖ) включают Irx4, Tbx20, IsIl, TnT, MLC2v и Tbx1. Фактор транскрипции eHAND экспрессируется в развивающемся ЛЖ и конотрункальной области и также имеет решающее значение для их развития. Др. генетические маркеры клеток первого поля сердца (ранний ЛЖ) включают Tbx5, Ncx2.5, TnT, MLC2V и HCN4.
Недавние исследования были посвящены регуляции групп генов, скоординированных в процессе развития. Один из механизмов заключается в экспрессии небольших некодирующих РНК, известных как микроРНК, каждая из которых регулирует экспрессию нескольких генов-мишеней. Др. механизм реализуется через модификации хроматина, каркаса ДНК, который действует как регулятор экспрессии генов. Ремоделирование хроматина, опосредованное такими факторами, как Brgl, Chd7, гистоновые деметилазы и метилтрансферазы, связано с пороками развития сердца.
б) Формирование сердечной петли. На ~22-24-й день сердечная трубка начинает изгибаться вентрально и вправо (см. рис. 1). Сердце — первый орган, который перестает подчиняться двусторонней симметрии раннего эмбриона. Асимметричная программа передачи сигналов, которая также влияет на положение легких, печени, селезенки и ЖКТ, определяет направление сердечной петли. Во время гаструляции, до начала формирования органов регулируется асимметричная экспрессия семейства генов sonic hedgehog (SHH*) и узловых генов (член семейства TGF-β) в латеральной мезодерме. Эти сигналы направленности создают градиент концентрации экспрессии критически важных сигнальных молекул между правой и левой сторонами эмбриона.
Такая асимметричная передача сигналов затем усиливается и распространяется через фактор транскрипции Pitx2, который экспрессируется в левой стороне ранней сердечной трубки, lefty 1 и LR динеин. Интересно, что мыши, у которых был инактивирован ген LR динеина, характеризуются случайной лево-правой (L-R) ориентацией сердца и внутренних ОБП, при этом в 50% случаев сердечная петля заворачивается вправо, а в 50% — влево. Др. потенциальные механизмы образования сердечной петли включают разную скорость роста миоцитов на выпуклой и вогнутой поверхности кривой, разную скорость запрограммированной гибели клеток (апоптоз) и механические силы, генерируемые внутри миокардиальных клеток на внутреннем и внешнем краях сворачивающегося сердца через актиновый цитоскелет.
P.S. * Sonic Hedgehog (SHH, «Сверхзвуковой ежик») — семейство генов и соответствующих им белков, управляющих эмбриональным развитием нервной системы и скелетной системы организма, а также выполняющих ряд др. физиол. функций.
Формирование петли приводит к перемещению будущего ЛЖ влево и созданию взаимосвязи с венозным синусом (будущие левое и правое предсердия), тогда как будущий ПЖ смещается вправо и образует сообщение с артериальным стволом (будущая аорта и ЛА). Такой тип развития объясняет относительно частое возникновение пороков сердца с удвоением выносящего тракта ПЖ и удвоением приточного тракта ЛЖ, а также крайне редкое удвоение выносящего тракта ЛЖ и удвоение приточного тракта ПЖ. При аномалиях формирования сердечной петли (situs inversus, гетеротаксии) высока частота серьезных пороков сердца, и обычно наблюдаются сопутствующие аномалии формирования легких и внутренних ОБП, включая отсутствие селезенки (аспления) или многочисленные маленькие селезенки (полиспления).
в) Формирование перегородок сердца. Когда формирование сердечной петли завершено, внешний вид органа становится похожим на зрелое сердце; внутренне структура напоминает единую трубку, хотя теперь она имеет несколько выпуклостей, приводящих к появлению примитивных камер. Общее предсердие (включающее правое и левое предсердия) соединяется с первичным желудочком (будущим ЛЖ) через АВ-канал. Примитивный желудочек соединяется с сердечной луковицей (будущим ПЖ) через бульбовентрикулярное отверстие. Дистальная часть сердечной луковицы соединяется с артериальным стволом через выносящий сегмент (конус).
Теперь сердечная трубка состоит из нескольких слоев миокарда и одного слоя эндокарда, разделенных сердечным гелем (бесклеточный внеклеточный матрикс, секретируемый миокардом). Формирование перегородок сердца начинается на ~26-й день с прорастания больших тканевых масс, эндокардиальных валиков, как в АВ-, так и в конотрункальном соединениях (см. рис. 1). Эти валики состоят из выступов внеклеточного матрикса (сердечного геля), которые, помимо своей роли в развитии, также выполняют физиологическую функцию в качестве примитивных сердечных клапанов. Эндокардиальные клетки дедифференцируются и мигрируют в сердечный гель в области эндокардиальных валиков, в конечном итоге становясь мезенхимальными клетками (эндотелиальномезенхимальная трансформация), которые в дальнейшем образуют часть АВ-клапанов.
Эндокард, вторичное поле сердца и нервный гребень также вносят вклад в формирование створок клапана. Помимо прямого вклада в клапанную ткань, эти клетки-предшественники также взаимодействуют друг с другом и с др. клетками сердца, управляя развитием сердечных клапанов.
Завершение формирования перегородки АВ-канала происходит за счет срастания эндокардиальных валиков. Большая часть ткани АВ-клапанов происходит из миокарда желудочков в процессе образования стенок желудочков. Поскольку этот процесс происходит асимметрично, кольцо трикуспидального клапана располагается ближе к верхушке сердца, чем кольцо митрального клапана. Физическое разделение этих двух клапанов приводит к образованию АВ-перегородки, отсутствие которой является основным распространенным дефектом у пациентов с пороками развития АВ-канала. Если процесс формирования полости желудочка не завершен, правый АВ-клапан не может нормально отделиться от миокарда желудочков, что является возможной причиной аномалии Эбштейна (Ebstein).
Формирование перегородки предсердий начинается на ~30-й день в/утробного развития первичной перегородки вниз к эндокардиальным валикам (см. рис. 1). При этом остается первичное отверстие. Затем эндокардиальные валики срастаются и вместе с завершенной первичной перегородкой разделяют АВ-канал на правый и левый сегменты. Вторичное отверстие, ostium secundum, появляется в задней части первичной перегородки — оно позволяет части венозного возврата плода в правое предсердие проходить в левое предсердие. Наконец, вторичная перегородка растет вниз, справа от первичной перегородки. Вторичное отверстие вместе с лоскутом первичной перегородки образует овальное окно, через которое кровь плода попадает из НПВ в левое предсердие.
Перегородка желудочков начинает формироваться на ~25-й день в/утробного развития с выступов эндокарда как в приточном (первичный желудочек), так и в выносящем (сердечная луковица) сегментах сердца. Выступы приточного отдела сливаются с бульбовентрикулярной перегородкой и разрастаются назад к нижнему эндокардиальному валику, где они дают начало приточной и трабекулярной частям МЖП. ДМЖП могут возникать в любой части развивающейся МЖП. Выносящий тракт или конотрункальная перегородка развивается из гребней сердечного геля, как и АВ-валики. Эти гребни сливаются, образуя спиральную перегородку, которая соединяет будущую ЛА с расположенным спереди и справа ПЖ, а будущую аорту — с расположенным сзади и слева ЛЖ.
Различия в росте клеток выносящей части перегородки приводят к удлинению гладкомышечного сегмента под легочным клапаном (конусом) — этот процесс обеспечивает разделение трикуспидального клапана и клапана ЛА. Напротив, исчезновение сегмента под аортальным клапаном приводит к анат. фиброзной целостности митрального и аортального клапанов. В просвете дистальной части выносящего тракта возникают местные тканевые утолщения (трункальные валики), которые позже заселяются мезенхимальными клетками, происходящими из нервного гребня и участвующими в формировании полулунных створок клапана ЛА и аортального клапана.
Нарушения этих процессов являются причиной конотрункальных дефектов и пороков развития дуги аорты (общий артериальный ствол, тетрада Фалло (Fallot), атрезия ЛА, удвоение выносящего тракта ПЖ, прерванная дуга аорты) — группы ВПС, часто связанных с делециями критической области ДиДжорджи (DiGeorge) хромосомы 22q11. Фактор транскрипции Tbx1 имеет значение как ген-кандидат, который м.б. причиной синдрома ДиДжорджи.
Несколько генов участвуют в формировании клапана, в т.ч. ген PTPN11, который кодирует тирозинфосфатазу Shp-2, а в мутированной форме является одним из генов, ответственных за развитие синдрома Нунан (Noonan), связанный со стенозом клапана ЛА, и NOTCH1, регулятор дифференцировки клеток, связанный с болезнями аортального клапана.
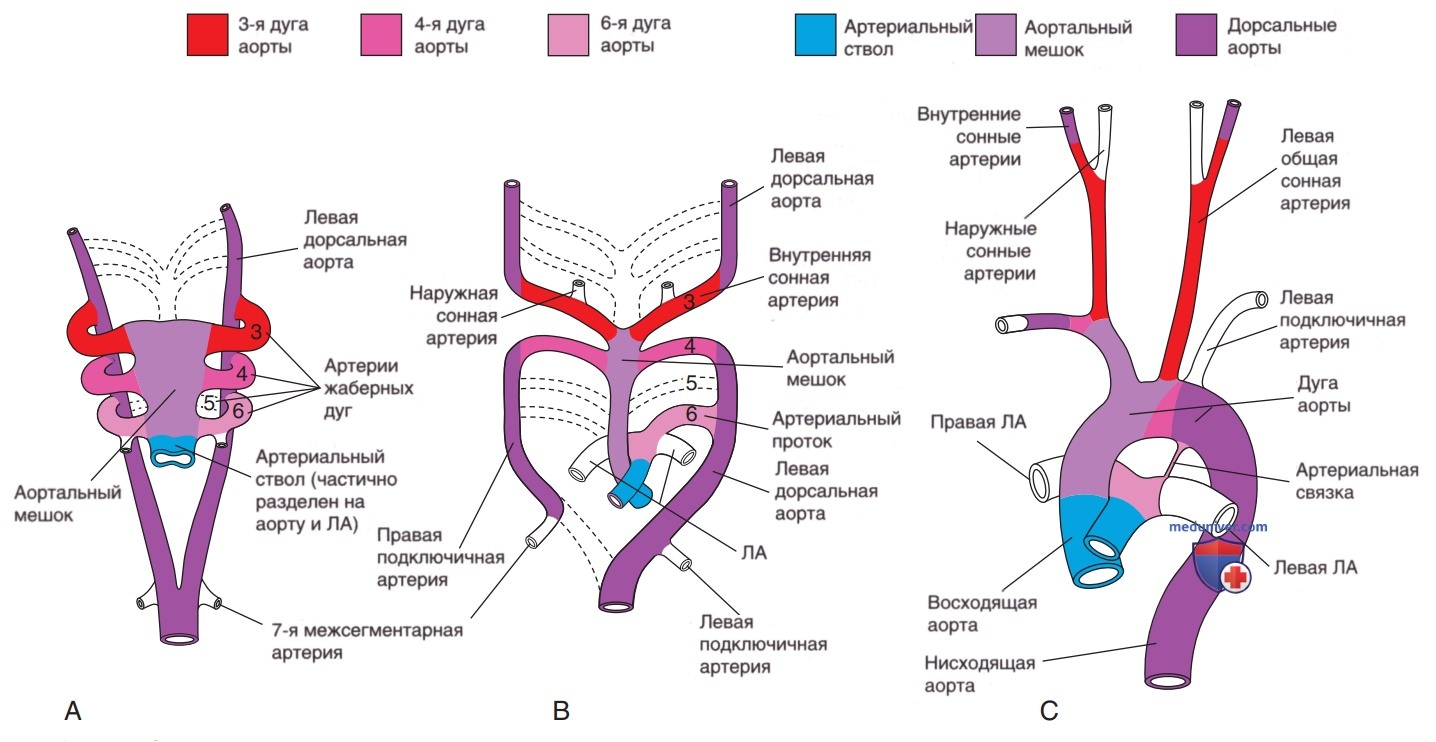
г) Развитие дуги аорты. Дуга аорты, сосуды головы и шеи, проксимальные ЛА и артериальный проток развиваются из аортального мешка, артериальных дуг и дорсальных аорт. При формировании прямой сердечной трубки дистальная часть выносящего тракта раздваивается на правую и левую 1-ю дуги аорты, которые соединяются с парными дорсальными аортами (рис. 2). Дорсальные аорты срастаются, образуя нисходящую аорту. Проксимальный отдел аорты от аортального клапана до левой сонной артерии развивается из аортального мешка. Первая и вторая дуги регрессируют к ~22-му дню в/утробного развития, причем 1-я дуга аорты дает начало верхнечелюстной артерии, а 2-я — стременной и подъязычной артериям.
Рисунок 2. Схематические иллюстрации изменений, которые происходят в результате трансформации артериального ствола, аортального мешка, аортальных дуг и дорсальных аорт в зрелое артериальное русло. Сосуды, которые не заштрихованы и не закрашены, не являются производными этих структур: А — дуги аорты на б-й неделе в/утробного развития; к этому моменту первые две пары дуг аорты уже практически исчезли; В — дуги аорты на 7-й неделе в/утробного развития; части дорсальных аорт и аортальных дуг, которые обычно исчезают, обозначены пунктирными линиями; С — артериальные сосуды ребенка в возрасте 6 мес.
Третья дуга участвует в формировании безымянной артерии, а также общей и внутренней сонных артерий. Правая 4-я дуга дает начало безымянной и правой подключичной артериям, а левая 4-я дуга участвует в формировании сегмента дуги аорты между левой сонной артерией и артериальным протоком. Пятая дуга не сохраняется как значимая структура в зрелом кровообращении. Шестая дуга соединяется с более дистальными ЛА, правая 6-я дуга дает начало проксимальной части правой ЛА, а левая 6-я дуга — артериальному протоку. Дуга аорты между артериальным протоком и левой подключичной артерией развивается из левой дорсальной аорты, тогда как дуга аорты дистальнее левой подключичной артерии формируется вследствие слияния правой и левой дорсальных аорт.
Аномалии развития парных дуг аорты становятся причиной образования правой дуги аорты, двойной дуги аорты и сосудистых колец.
д) Дифференцировка сердца. Процесс, благодаря которому тотипотентные клетки раннего эмбриона становятся коммитированными в определенные клеточные линии, называется дифференцировкой. Прекардиальные мезодермальные клетки дифференцируются в зрелые клетки сердечной мышцы с соответствующим набором специфичных для сердца сократительных элементов, регулирующих белков, рецепторов и ионных каналов. Экспрессия сократительного белка миозина происходит на ранней стадии развития сердца, даже до слияния двусторонних зачатков сердца. Дифференцировка в этих ранних мезодермальных клетках регулируется сигналами от передней эндодермы — этот процесс известен как индукция. Несколько предполагаемых ранних сигнальных молекул включают фактор роста фибробластов, активин и инсулин.
Сигнальные молекулы взаимодействуют с рецепторами на поверхности клетки; эти рецепторы активируют вторичные мессенджеры, которые, в свою очередь, активируют специфические факторы ядерной транскрипции (GATA-4, MEF2, Nkx, bHLH и семейство рецепторов ретиноевой кислоты), которые индуцируют экспрессию продуктов специфических генов для регулирования дифференцировки сердца. Ряд первичных заболеваний сердечной мышцы, кардиомиопатий, м.б. связан с дефектами некоторых из этих сигнальных молекул.
Процессы развития специфичны для вида камеры сердца. На ранних стадиях развития миоциты желудочков экспрессируют как желудочковые, так и предсердные изоформы нескольких белков, таких как предсердный натрийуретический пептид (ANP; англ. atrial natriuretic peptide) и легкая цепь миозина (MLC; англ. myosin light chain). Зрелые миоциты желудочков не экспрессируют ANP и экспрессируют только специфичную для желудочков изоформу MLC 2v, тогда как зрелые миоциты предсердий экспрессируют ANP и специфичную для предсердий изоформу MLC 2а.
СН, перегрузка объемом и гипертрофия вследствие перегрузки давлением связаны с рекапитуляцией фенотипов фетальных клеток, при которых зрелые миоциты повторно начинают экспрессировать фетальные белки. Поскольку разные изоформы имеют разное сократительное поведение (быстрая или медленная активация, высокая или низкая активность аденозинтрифосфатазы), экспрессия разных изоформ может иметь важные функциональные последствия.
е) Изменения сердечной функции в процессе развития. В процессе развития состав миокарда претерпевает глубокие изменения, в результате которых увеличивается количество и размер миоцитов. Во время в/утробной жизни этот процесс включает деление миоцитов (гиперплазию), тогда как после первых нескольких постнатальных недель последующий рост сердца происходит в основном за счет увеличения размеров миоцитов (гипертрофия). Сами миоциты меняют форму с круглой на цилиндрическую, возрастает доля миофибрилл (содержащих сократительный аппарат), и миофибриллы приобретают более упорядоченную ориентацию.
Плазматическая мембрана (известная как сарколемма в миоцитах) — это место локализации ионных каналов и трансмембранных рецепторов, которые регулируют обмен хим. информацией между поверхностью клетки и ее внутренней средой. Потоки ионов через эти каналы управляют процессами деполяризации и реполяризации. Описаны изменения в процессе развития для натрий-калиевого насоса, натрий-водородного обменника и потенциал-зависимых кальциевых каналов. По мере созревания миоцита сарколемма разрастается по направлению к внутренней части клетки (система Т-трубочек), что резко увеличивает площадь ее поверхности и способствует быстрой активации миоцита. Регуляция мембранных α- и β-адренорецепторов в процессе развития увеличивает способность СНС контролировать функцию сердца по мере его созревания.
Саркоплазматический ретикулум, система канальцев, окружающих миофибриллы, контролирует в/клеточную концентрацию кальция. Группа насосов регулирует высвобождение кальция в миофибриллы для инициирования сокращения (рианодин-чувствительный кальциевый канал) и захват кальция для инициирования процесса расслабления (аденозинтрифосфат-зависимый кальциевый насос саркоплазматического ретикулума). Эта система транспорта кальция в саркоплазматический ретикулум менее развита в незрелом сердце, которое, т.о., больше зависит от транспорта кальция извне клетки для сокращения. В зрелом сердце большая часть кальция, активирующего сокращение, поступает из саркоплазматического ретикулума. Этот феномен процесса развития может объяснить чувствительность сердца младенца к блокаторам сарколеммальных кальциевых каналов, таким как верапамил, который может привести к заметному снижению сократимости.
Основные сократительные белки (миозин, актин, тропомиозин и тропонин) организованы в функциональную единицу сердечного сокращения — саркомер. У каждого сократительного белка есть несколько изоформ, которые по-разному экспрессируются в зависимости от местоположения (предсердие или желудочек) и стадии развития (эмбрион, плод, новорожденный, взрослый).
Изменения в структуре миокарда и биохимии миоцитов приводят к легко поддающимся количественной оценке различиям в сердечной функции в процессе развития. Функция сердца плода менее чувствительна к изменениям как преднагрузки (объем наполнения), так и постнагрузки (системное сопротивление). Самым эффективным средством усиления функции желудочков у плода является увеличение ЧСС. После рождения и по мере дальнейшего созревания преднагрузка и постнагрузка играют все более важную роль в регулировании сердечной функции. Скорость расслабления сердечной мышцы также зависит от фазы развития. Снижение способности незрелого кальциевого насоса саркоплазматического ретикулума удалять кальций из сократительного аппарата проявляется уменьшением возможностей сердца плода усиливать расслабление в ответ на симпатическую стимуляцию.
Видео эмбриогенез (развитие) сердца и его аномалии