С момента рождения младенец подвергается воздействию микробов, находящихся на теле матери и в окружающей среде. Микроорганизмы быстро образуют скопления на открытых участках тела, включая кожу и энтеральный тракт. Микробные сообщества, называемые микробиотой, влияют на краткосрочную и долгосрочную физиологию, включая иммунологическое и метаболическое развитие и функции организма. В совокупности количество ассоциированных с организмом бактериальных клеток оценивается в 10 раз больше, чем количество человеческих клеток в организме.
Совокупность микробов, включая их гены и взаимодействия с окружающей средой, составляет микробиом, а количество микробных генов в микробиоме человека превышает количество человеческих генов как минимум в 100 раз, вместе составляя макроорганизм с неразрывной коллективной физиологией. Текущие данные показывают, что микробиом эволюционирует на протяжении жизни, влияя на здоровье и развитие заболеваний.
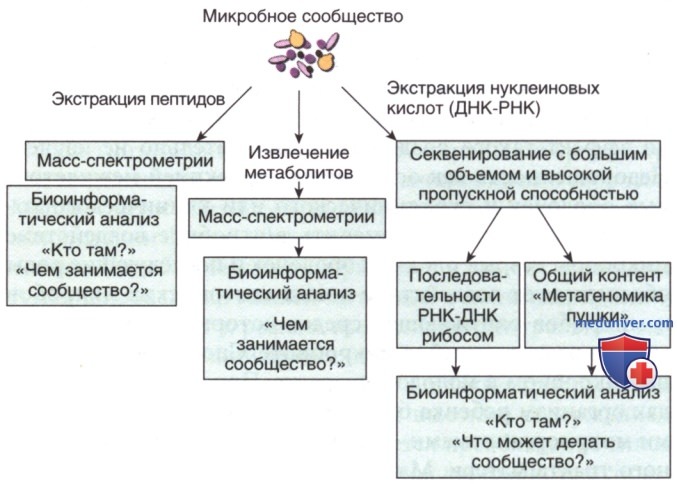
а) Оценка микробиома. Предыдущие знания о микробах на теле человека и вокруг него основывались на конкретных методах культивирования организмов. Молекулярные технологии произвели революцию в идентификации плохо культивируемых микробов, редких микробов и микробов в сложных сообществах, например, связанных с человеческим телом (рис. 1).
Рисунок 1. Общие молекулярные методологии для идентификации компонентов и функций сложных микробных сообществ
Развитие технологии ПЦР и доступность современных методов секвенирования нуклеиновых кислот повысили чувствительность обнаружения многих организмов и привели к открытию новых разновидностей. Современные технологии секвенирования, называемые платформами секвенирования следующего поколения, позволяют выполнять секвенирование большого объема и глубины с миллионами последовательностей, полученными из одного биологического образца.
Три основных подхода используют секвенирование следующего поколения для понимания состава, разнообразия и активности микробиома: (1) секвенирование видоспецифичных областей генома — треки, кодирующие рибосомную РНК, и межгенные области, называемые метагеномикой, (2) полное секвенирование ДНК из образцов (фекалии, слюна) и сборка фрагментов последовательности в большие фрагменты генома, называемые «пушечной» метагеномикой, и (3) секвенирование транскрипта РНК для расшифровки состава и, в качестве суррогата функциональной активности, транскрипционной активности микробиома — метатранскриптомики. Огромные вычислительные мощности и новые инструменты биоинформатики позволили анализировать и сравнивать большие наборы данных, полученные с помощью этих методов.
Кроме того, разработаны 2 дополнительных подхода к измерению фенотипа микробиома. Во-первых, крупномасштабные измерения пептидного состава микробиоты, называемые протеомикой, чаще используются для описания активности образца микробиома, поскольку пептиды предоставляют информацию о составе и функции микробиома. Во-вторых, в дополнительном подходе, называемом метаболомикой, метаболиты, полученные из микробиома, измеряются с использованием расширенных методов газовой хроматографии и масс-спектрометрии.
Вместе протеомика и метаболомика лучше описывают активность микробиома, чем подходы к нуклеотидному секвенированию. Однако на данном этапе они обеспечивают меньшую глубину разрешения и специфичность по сравнению с составом и фенотипом микробиома. Эти методы позволяют исследовать микробиом, но еще не заменяют культивирование микробов во многих клинических случаях. Культивирование организмов по-прежнему наиболее практичный способ дифференциации потенциальных патогенов от более доброкачественных видов и предоставления клинически действенной информации (чувствительность к ряду АБ).
б) Раннее развитие микробиома в детстве. Новые исследования показывают, что плацента и плод подвергаются воздействию микробов в/утробно, но эффект такого воздействия окончательно не изучен. Недоношенность как осложнение инфекцией межклеточных оболочек и субклинического или клинического хориоамнионита может изменить в/утробное воздействие микробов. Разрыв плодных оболочек и последующие роды обеспечивают воздействие новых материнских микробов и микробов окружающей среды, которые займут общие места в развивающейся микробиоте. Способ родов влияет на микробиом в молодом возрасте. При естественных родах организм ребенка быстро колонизируются кишечными микроорганизмами — отражение состояния вагинального тракта матери.
Младенцы, родившиеся посредством кесарева сечения, колонизируются микроорганизмами, отражающими состояние кожи и ротовой полости матери (стафилококки и стрептококки), а также микроорганизмами из окружающей среды. У доношенного ребенка, родившегося естественным путем, первые кишечные микробы («новаторские организмы»), включают Escherichia и др. Enterobacteriaceae, Bacteroides и Parabacteroides. Исключительно грудное вскармливание приводит к высокому уровню бифидобактерий и лактобактерий в течение недели после начала кормления. Эти пробиотические организмы обладают уникальной способностью исключать потенциальные патогены из колонизации путем связывания питательных веществ и выработки антимикробных факторов, одновременно стимулируя эпителий кишечника для уплотнения клеточных контактов и экспрессии антимикробных пептидов.
Недоношенный ребенок с большей вероятностью рождается путем кесарева сечения: его организм в большей степени заселен кожными микроорганизмами (коагулазонегативные стафилококки). Аналогичная картина наблюдается у доношенных детей, родившихся путем кесарева сечения. Однако недоношенный ребенок не всегда проходит те же стадии расширения и диверсификации микробиома в течение 1 нед-1 мес жизни, что и доношенный ребенок. Факторы, связанные с отсроченным созреванием: отсроченное или ограниченное энтеральное кормление, нормальное воздействие окружающей среды на домашнюю среду и медицинские вмешательства (АБТ).
Наиболее значительный сдвиг в микробиоте кишечника происходит после отлучения от груди и введения прикорма. По мере того, как младенец переходит от грудного молока к твердой пище, содержащей сложные полисахариды растительного происхождения, микробиота постепенно преобразуется в более зрелую композицию, напоминающую микробиоту взрослых. В то же время метаболический потенциал микробиома изменяется, чтобы приспособиться к изменяющейся диете: микробиом новорожденного обогащается генами фосфотрансферазной системы (PTS). К 4 мес микробиом смещается в сторону увеличения количества генов - переносчиков лактозы, что отражает потребление молока, и к 12 мес происходит дальнейший переход к большему количеству генов, ответственных за работу ферментов-переносчиков β-глюкозидов, необходимых для расщепления сложных углеводов.
Созревание детского микробиома после первых лет жизни до взрослого возраста менее изучено, и необходимы дополнительные исследования с большой выборкой, чтобы полностью понять стадии созревания и достижения сходства со зрелым, здоровым взрослым состоянием.
Микробиота полости рта новорожденных ассоциирована с микробиотой матери. У младенцев, рожденных естественным путем, преобладают Lactobacillus, Prevotella и Sneathia, а у детей, рожденных после кесарева сечения, больше материнских кожных микроорганизмов (Staphylococcus, Corynebacterium и Propionobacterium). В течение 1 дня жизни в полости рта преобладают Firmucutes, стрептококк и стафилококк. У младенцев, находящихся на искусственном вскармливании, наблюдается больше Bacteroidetes, а у детей, находящихся на грудном вскармливании, — больше бактерий типа Proteobacteria и Actinobacteria.
С прорезыванием первых зубов формируются новые экологические ниши, способствующие развитию микробных сообществ. Ранее считалось, что кариесогенные бактерии (Streptococcus mutans) приобретаются после прорезывания зубов. Однако недавние данные демонстрируют присутствие этих организмов до прорезывания зубов в резервуаре мягких тканей, подчеркивая важность хорошего ухода за полостью рта у младенцев даже до первичного прорезывания зубов.
К 3 годам микробиом ротовой полости и слюны у детей становится сложным, но менее разнообразным, чем микробиом взрослого человека. Состав микробиоты ротовой полости при полностью сформированных зубных рядах взрослого человека насчитывает 1000 видов бактерий.
Даже с учетом здоровья полости рта, разнообразие микрофлоры значительно: оно меняется с развитием заболеваний полости рта (например, пародонтит). Неизвестно, как микробиота развивается между дошкольным и взрослым возрастом. Установка и снятие стоматологической аппаратуры для ортодонтии в детстве может вызвать значительные изменения в микробиоме ротовой полости. В течение первого года жизни микробиом кожи младенца увеличивается в разнообразии, видовом богатстве и однородности. Кожа ребенка младшего возраста относительно недифференцирована между участками тела, с большим количеством общих микроорганизмов на разных участках тела (руки, лоб и ягодицы). У детей старшего возраста микробные сообщества на каждом участке тела дифференцированы.
Кожа младенцев преимущественно заселена Firmicutes (Streptococcaceae и Staphylococcaceae, актинобактерии, протеобактерии и Bacteroidetes). Микробиом кожи взрослого человека демонстрирует высокую степень георазнообразия — различия в составе в зависимости от места и физиологии. Наиболее значимые различия микробиома наблюдаются на участках сухой и влажной кожи. Связь между развитием кожи в детстве и созреванием микробиома кожи — предмет текущих исследований. Социальная структура и семейные взаимодействия играют важную роль в развитии микробиома в раннем возрасте.
Кормление грудным молоком обеспечивает микробиологическую связь между матерью и младенцем: передачу пробиотикоподобных организмов (лактобациллы и бифидобактерии). Каждый из этих микроорганизмов имеет защитные эффекты, включая защиту от диарейных заболеваний и атопии. Педиатры осведомлены о рисках инфекционных заболеваний и преимуществах посещения детских садов. Пример: общие штаммы пневмококка, вызывающие средний отит и вспышки РСВ-инфекции и ассоциации со снижением атопии, аллергии и БА. Семейные контакты — риск MRSA и последующего заболевания. Исследования также демонстрируют передачу частей человеческого микробиома домашним животным (собакам и кошкам).
Например, у семьи определяются одни и те же штаммы кишечной палочки, которые вызывают ИМП у одного из членов семьи. Могут быть различия в микробиоте полости рта среди младенцев, получавших и не получавших соску. В сельской местности общность микробиома распространяется на домашний скот, помещения и поверхности, членов семьи. Развитие микробиома в детстве при взаимодействии с окружающей средой — сложный процесс, который до сих пор исследуется.
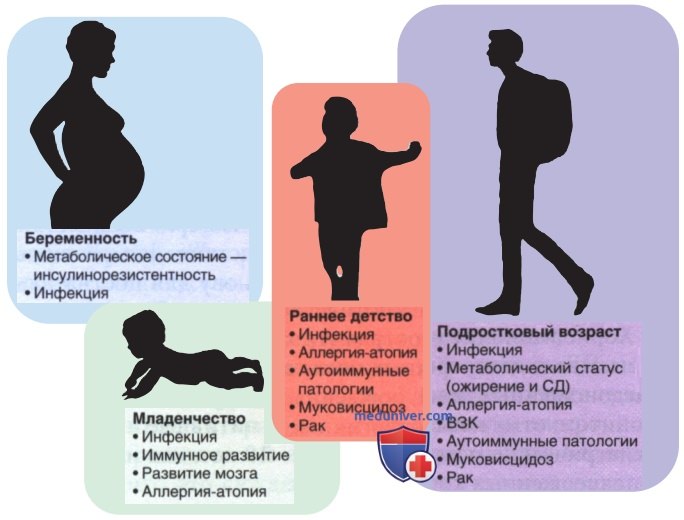
в) Микробиом и физиологическое развитие. Микробиом играет важную роль в физиологии млекопитающих (рис. 2). Микробиота участвует в развитии энтерального тракта, ДП, иммунной системы, гематологической системы, метаболико-эндокринной системы и НС. Детали того, как микробиом способствует физиологическим процессам человеческого организма, до сих пор исследуются. Однако моделирование роли микробиома в развитии органов и систем млекопитающих указывает, что функции микроорганизмов варьируют в зависимости от вида.
Рисунок 2. Физиологические и патологические роли микробиома, связанные с педиатрией. Микробиом человека влияет на здоровье и развитие ребенка от беременности до зрелого возраста, включая инфекции и процессы, не связанные с инфекциями
1. Микробном и метаболизм. Вскоре после перехода во внешний мир энтеральный тракт млекопитающих колонизируется, и взаимодействие первых микробов в энтеральном тракте стимулирует развитие слизистой оболочки кишечника. В моделях новорожденных и молодых животных отсроченная или отсутствующая колонизация кишечника приводит к неполному развитию эпителия, уплощению кишечных крипт, потере сосудистой сети и серьезному снижению ферментативной функции, включая ЩФ и глюкозидазы. Кишечная микробиота играет большую роль в физиологии кишечного тракта. Она стимулирует развитие слизистой оболочки и системного иммунитета, развитие и регенерацию эпителия и эндотелия, созревание клеток и поддержание метаболизма. Метаболизм включает: (1) переваривание в целом неперевариваемых растительных полисахаридов; (2) производство витаминов и кофакторов; (3) метаболизм ксенобиотиков; (4) стимуляцию местного и системного метаболизма, включая накопление липидов.
У гнотобионтов (животных, выращенных в стерильных условиях без контакта со внешней средой), лишенных кишечной микробиоты, наблюдается ограниченное количество питательных веществ и фенотип, неспособный к развитию. Мыши-гнотобионты, рожденные в стерильной среде, служат моделью для понимания роли микробиома в здоровье. Мыши-гнотобионты гуманизируются путем селективной колонизации фекальными микробными сообществами человека. Подобно введению прикорма у людей, кормление гуманизированных мышей питанием с полисахаридами и без них приводит к резким изменениям центральных метаболитов. У гуманизированных мышей, переведенных с богатой полисахаридами диеты с низким содержанием жиров на более западную диету с высоким содержанием жиров и моносахаридов, наблюдается преобладание типов Actinobacteria и Firmicutes в кишечной микробиоте с соразмерным сокращением Bacteroidetes. Аналогичные изменения — увеличение количества Firmicutes и уменьшение Bacteroidetes — наблюдаются при ожирении у человека.
У людей существуют общие закономерности состава зрелой кишечной микробиоты и ее прогнозируемой функции. Секвенирование фекальных микробов взрослых людей из разных стран выявило 3 общих паттерна состава микробного сообщества — биотипы. Bacteroides, Prevotella и Ruminococcusin — 3 доминирующих рода микроорганизмов. Биотипы различаются у людей с разных континентов, включая Северную Америку, Европу и Азию, что отражает культурные и диетические различия. Микробиом младенца значительно варьирует; зрелые устойчивые биотипы формируются в раннем послеродовом периоде и в младенчестве. Описаны биотипы грудного молока и искусственного вскармливания с заметным обогащением кишечными грамотрицательными бактериями (Е. coli и анаэробные Clostridia spp.) среди младенцев, находящихся на искусственном вскармливании. Влагалищные биотипы молодых и стареющих женщин различаются в зависимости от возраста, расы и этнической принадлежности.
2. Микробиом, воспаление и иммунитет. Микроорганизмы, из которых состоит микробиом, значимы для раннего иммунопрограммирования, развития иммунной толерантности и общего поддержания иммунных установок. Клетки вырабатывают множество рецепторов для распознавания микробных лигандов в процессе, называемом распознаванием образов. Микробы преднамеренно и непреднамеренно стимулируют эти клеточные рецепторы для активации и подавления воспалительных путей. Классические примеры таких регуляторных взаимодействий — пептидогликаны на бактериях, связывающиеся с Toll-подобным рецептором 2 (TLR-2, в комплексе с TLR-3 и TLR-6; англ. Toll-like receptor), липополисахариды грамотрицательных бактерий, связывающиеся с TLR-4, и глюканы грибов, связывающиеся с рецепторами дектина. Результаты этих взаимодействий с рецепторами — производство хемокинов и цитокинов, дифференцировка и развитие клеток, изменение метаболизма и стимуляция программ гибели и выживания клеток.
Все зависит от типа клетки, ее состояния и величины стимуляции. Стимуляция систем распознавания микробов важна для развития: у животных, выращенных в стерильной среде, снижен врожденный иммунный ответ. Например, не вырабатываются антимикробные пептиды на поверхности слизистых оболочек, нарушается регуляция провоспалительных и иммунологических реакций толерантности и снижаются популяции Т- и В- клеток. После восстановления нормальной колонизации ЖКТ через несколько недель после выхода из стерильной зоны у животных сохраняются долгосрочные аберрантные цитокиновые реакции с гиперактивными провоспалительными реакциями на стимулы. Это стойкие последствия изменения раннего приобретения микробов. Различные модели колонизации в раннем возрасте также коррелируют с долгосрочным иммунным развитием.
В скандинавском исследовании дети с персистирующей колонизацией Е. coli в раннем возрасте имели более высокие уровни устойчивых В-клеток памяти (CD3+CD20+CD27+) к 1,5 годам, чем дети с более низким уровнем колонизации Е. coli, даже несмотря на обильную колонизацию прототипическими пробиотическими бактериями Lactobacillus.
3. Микробиомно-нейробиологические связи. Исследования демонстрируют: ось кишечник-мозг изменяется в зависимости от состава и активности кишечного микробиома. Исследования на животных моделях показали, что микробиом изменяет систему гипоталамус-гипофиз-надпочечники.
Мыши гнотобионты демонстрируют чрезмерно тревожно-стрессовое поведение, сопровождающееся повышенными уровнями кортикостерона и АКТГ, по сравнению с мышами, колонизированными обычным способом, без патогенов. Нейропластичность, включая нейрогенез и активацию микроглии, регулируется микробиотой. Функциональная МРТ показала, что прием 5 штаммов пробиотических бактерий изменяет активность мозга у людей: это приводит к снижению реакции мозга на задачи эмоционального внимания в сенсорных и эмоциональных областях мозга.
Механизм, лежащий в основе этих изменений, окончательно не изучен: предполагается, что tractus solitarius и блуждающий нерв опосредуют связь ЖКТ и ГМ. Другой механизм, с помощью которого кишечный микробиом может изменять активность мозга, — производимые им метаболиты. Введение в ферментированное молоко пробиотических организмов (Bifidobacterium animalis subsp. Lactis) у монозиготных человеческих близнецов и мышей не изменило кардинально состав кишечного микробиома, но скорректировали его транскрипционные профили с переходом к усилению ферментации углеводов в жирные кислоты, которые уменьшают негативные эмоциональные реакции у людей.
г) Роль микробиома при заболеваниях. Исследования показывают, что некоторые микробные сообщества действуют сообща, оказывая негативное влияние на здоровье. Др. микроорганизмы отвечают за восстановление защитных механизмов или устойчивость к болезням. Некоторые примеры концепции измененных микробных сообществ, также называемой дисбиозом, представлены в следующих разделах.
1. Микробиом при преждевременных родах. Этиология преждевременных родов многофакторная, но воспалительные состояния(субклинические и клинически явные инфекции матери и плода) провоцируют их начало. Профили воспалительных биомаркеров подчеркивают этот момент: у женщин с преждевременными родами повышен уровень ангиотензина, ИЛ-8 и рецептора ФНО, наряду с расовыми изменениями дополнительных цитокинов и хемокинов. У женщин, перенесших преждевременные роды, увеличена колонизация влагалища Gardnerella spp. и Lactobacillus crispatus. Разнообразие микробиоты заднего свода влагалища у женщин, перенесших преждевременные роды, ниже, чем у женщин, родивших в срок.
Метаанализ раннего лечения вагиноза клиндамицином до 22 нед беременности продемонстрировал снижение числа спонтанных преждевременных родов на сроке <37 нед. Это согласуется с корреляцией между дисбактериозом микробиоты, связанной с беременностью, и преждевременными родами.
Считается, что амниотический пузырь и плод стерильны до разрыва плодных оболочек и рождения. Однако в нескольких отчетах обнаружены доказательства наличия бактериальной ДНК в меконии с 2 преобладающими типами мекония независимо от способа родоразрешение: (1) с преобладанием Enterobacteriaceae и (2) с преобладанием Leuconostocaceae, Enterococcaceae и Streptococcaceae. Кроме того, данные показывают, что в околоплодных водах при субклиническом и клинически очевидном хориоамнионите присутствуют микробы вагинального происхождения, включая плохо или не культивируемые организмы Mycoplasma spp., Ureaplasma spp., Bacteroides spp., Fusobacterium, Sneathia amangupinegens и Leptotrichia amnionii.
Существует корреляция между количеством интраамниотических организмов и степенью недоношенности. Микробная инвазия в амниотическое пространство приводит к индукции воспалительных путей через рецепторы распознавания микробных паттернов врожденного иммунитета (TLR). Результат — стимуляция родов и физиологический стресс у плода и матери. Воздействие микробных факторов имеет последствия для развития легких и кишечника, закладывая основу для постнатальной патологии, в т. ч. некротического энтероколита.
Хориоамнионит — острая угроза матери и плоду, но он не вызывает долгосрочных последствий для развития НС. У недоношенных детей, рожденных от женщин с хориоамнионитом, отмечены аналогичные когнитивные и нейропсихиатрические исходы в возрасте <18 лет, как и у детей, не подверженных хориоамниониту.
2. Изменения микробиома при некротическом энтероколите. Некротический энтероколит — тяжелое заболевание кишечника новорожденных, которое поражает сильно недоношенных детей с МТ <1500 г при рождении. Патологические стадии при некротическом энтероколите: воспаление кишечника с потерей барьерной функции, микробная инвазия в кишечник и некроз пораженных тканей кишечника. Исследования выявили определенные организмы как причину некротического энтероколита в серии случаев, но ни одна из предложенных специфических этиологий не оказалась общей для всех случаев заболевания. Многие микроорганизмы колонизировали кишечник после перенесенной патологии.
В патогенезе некротического энтероколита рассматривают модель дисбактериоза кишечного микробиома в раннем возрасте. Эпидемиологические исследования младенцев с ОНМТ при рождении продемонстрировали связь между приемом цефалоспоринов и продолжительностью воздействия АБ с развитием некротического энтероколита. Это согласуется с идеей, что изменения в микробиоте предрасполагают к некротическому энтероколиту или провоцируют его.
Исследования демонстрируют снижение разнообразия микробиоты до и во время некротического энтероколита. Микробиота кишечника при некротическом энтероколите во время клинических симптомов напоминает микробиоту за 72 ч до появления симптомов, но не микробиоту за 1 нед до появления симптомов. Это позволяет предположить, что изменение микробиоты кишечника начинается задолго до появления некротического энтероколита. Некоторые различия в ранней колонизации после рождения могут предвещать повышенный риск некротического энтероколита.
3. Микробиом и аллергические расстройства. Учитывая роль микробиома в развитии и модуляции врожденных и адаптивных иммунных ответов, значительный интерес вызывает его роль в развитии и обострении аллергических состояний (например, атопический дерматит). Микробиом кожи изучен до, во время и после лечения обострений атопического дерматита. Вспышки приводят к потере разнообразия бактерий на пораженной области, а лечение вносит новое разнообразие. Staphylococus aureus и S. epidermidis увеличиваются до и во время атопических обострений, a Streptococcus и Corynebacterium spp. увеличиваются непосредственно до и во время клинического улучшения.
У мышей пероральное лечение невсасывающимися АБ увеличивает уровень сывороточного иммуноглобулина (IgE), усиливает клинические симптомы (зуд) и вызывает признаки атопии. Эти данные свидетельствуют о том, что на атопический дерматит влияют местный микробиом кожи и более удаленные микробиомы, например микробиота ЖКТ. Этим объясняется тот факт, что введение пероральных пробиотиков (Lactobacillus spp.) уменьшает выраженность симптомов атопического дерматита с сопутствующим сдвигом баланса Т-хелперов (Th1/Th2) и повышением уровня интерферона-γ (часть иммунной толерантности).
Аллергические заболевания часто поражают ДП, а инфекции ассоциируются с их обострениями. Ранее считалось, что НДП стерильны, но исследования микробиома ДП у здоровых детей и взрослых, страдающих БА, показывают, что это суждение ошибочно. У здоровых людей при бронхоскопическом исследовании и цитологических соскобах наблюдается значительное разнообразие микрофлоры ДП. Измерение микробиоты НДП у здоровых детей и детей с БА указывает на существенные различия. Прошлые исследования на основе культур показывают, что колонизация респираторного дерева новорожденных в раннем возрасте Haemophilus influenzae, Moraxella catarrhalis и Streptococcus pneumoniae связана с повышенным риском детской БА. Эти же организмы связаны с обострениями БА.
В мышиной модели колонизация носоглотки новорожденных Н. influenzae в раннем возрасте приводит к снижению регуляторных Т-клеток ДП. У колонизированных животных наблюдается повышенная гиперчувствительность ДП после сенсибилизации аллергеном и ингаляционного заражения. Mycoplasma pneumoniae рассматрвиается в качестве основного бактериального индуктора обострений детской БА при выявлении инфекции.
Использование независимых от культуры измерений состава микробиоты нижних ДП (рис. 1) показало: у детей с БА более высокий уровень протеобактерий (Н. influenzae, Firmicutes, Staphylococcus и Streptococcus spp.). У здоровых детей чаще, чем у асматиков, определялись Bacteroidetes в НДП, особенно Prevotella spp. (групп анаэробных бактерий). Наличие у ЗЛ анаэробной популяции бактерий в НДП неожиданно: среда с высоким содержанием кислорода ранее считалась токсичной для анаэробов. Это исследование показывает, что среда ДП значительно отличается от ранее предполагавшейся, и необходимо изучить потенциально защитные свойства местной микробиоты, связанной со здоровьем.
4. Микробиом дыхательных путей при муковисцидозе. Муковисцидоз характеризуется прогрессирующим заболеванием ДП и воспалением с обострениями, сопровождающимися нарушением функции легких. У пациентов с муковисцидозом наблюдается возрастное изменение колонизации НДП, которое начинается в раннем детстве с S. aureus и Н. influenzae и смещается в сторону более естественных организмов с множественной лекарственной устойчивостью, включая резистентные к лечению бактерии Pseudomonas aeruginosa и Burkholderia cepacia complex. Независимый от микробной культуры молекулярный анализ микробиоты легких при муковисцидозе выявил более сложные микробные сообщества, чем ожидалось ранее, и продемонстрировал связь между возрастом пациента и тяжестью заболевания. В дополнение к присутствию множества ранее неожиданных микроорганизмов ДП (анаэробы и микобактерии) тяжесть заболевания обратно пропорциональна разнообразию микробного сообщества НДП.
При этом менее распространенное заболевание связано с большим видовым богатством и однородностью микробиоты. Напротив, потеря разнообразия, в т.ч. переход от менее сложных микробных сообществ к сообществам, в которых доминирует Р. aeruginosa, сильно коррелирует с тяжестью заболевания, а уровни Н. influenzae отрицательно коррелируют с серьезностью заболевания. Хотя АБ снижают скорость прогрессирующего ухудшения функции легких, они также уменьшают разнообразие микробного сообщества. Нарушается баланс между разнообразной микробиотой и доминированием отдельных организмов (Р. aeruginosa).
5. Микробиом при антибиотик-ассоциированной диарее и колите, вызванном Clostridium difficile. Лечение пероральными и парентеральными АБ приводит к быстрому и значительному изменению микробиоты кишечника. У здоровых участников исследования, принимающих ципрофлоксацин, наблюдаются неблагоприятные, но индивидуализированные изменения микробиома в ответ на АБ. Значительно сокращается количество бактерий за пределами ожидаемого спектра, что подчеркивает взаимозависимость членов микробного сообщества друг от друга для их стабильности в сообществе в целом. Кроме того, реакция на ципрофлоксацин среди участников варьировала в зависимости от человека: это свидетельство разной степени стабильности микробиоты и устойчивости к стрессу (прием АБ). Микробное сообщество в большей степени восстановилось в течение 4 нед после завершения курса АБ.
Некоторые АБ, например (ампициллин + клавуланат), вызывают диарею. Диарея ассоциирована с потерей Clostridium и Bacteroides, которые играют важную роль в производстве короткоцепочечных жирных кислот (SCFA; англ. Short-Chain Fatty Acids) и метаболизме неперевариваемых углеводов. Их совместная потеря снижает метаболическую целостность кишечного эпителия, который использует SCFAs для получения энергии, в результате чего образуется высокоосмотическая среда, в которой жидкость втягивается в просвет кишечника. Диарея, связанная с АБ, может быть результатом этих комбинированных эффектов. У >30% младенцев колонизация С. difficile начинается с 1 мес жизни и продолжается до 6 мес. К 1 году колонизация составляет 15-70%, а затем снижается до взрослого возраста, когда носительство оценивается в <3%.
С. difficile обнаружена в микробиоте влагалища беременных женщин, но естественные роды не связаны с повышенной частотой колонизации кишечника новорожденных С. difficile. При естественных родах и родах путем кесарева сечения частота колонизации составляла 30 и 37% соответственно. Clostridium-ассоциированная диарея приводит к 35-45 госпитализациям на 10 000 педиатрических госпитализаций среди детей 1-9 лет.
Исследования еще не определили естественное состояние кишечного микробиома до, во время и после Clostridium-ассоциированной диареи у детей. Молекулярные исследования кишечной микробиоты у взрослых предоставляют некоторые детали последствий Clostridium-ассоциированной диареи для кишечной микробиоты. Исследования, в которых использовалось глубокое секвенирование стула людей с Clostridium-ассоциированной диареей и колонизацией С. difficile без заболевания, выявили истощение определенных родов бактерий, сопровождающее присутствие колонизации С. difficile. К этим родам относятся Blautia, Pseudobutyri vibrio, Roseburia, Faecali bacterium, Anaerostipes, Subdoligranulum, Ruminococcus, Streptococcus, Dorea и Coprococcus.
Причинно-следственная связь изменений микробиома и событий, запускающих переход от колонизации к симптоматическому заболеванию, неизвестна, но связана с истощением конкурентных видов С. difficile. Подобно исследованиям диареи, связанной с АБ, эти исследования демонстрируют снижение количества производящих бутират Clostridium spp. Бутират — источник энергии для кишечного эпителия и поддержания его целостности. Для лечения Clostridium-ассоциированной диареи применялись АБ метронидазол и ванкомицин, но традиционное лечение не устраняет рецидивирующее заболевание полностью. Для решения этой проблемы применяется трансплантация фекалий или введение фекалий здоровых доноров реципиентам с Clostridium-ассоциированной диареей. Это экономически эффективное лечение, которое превосходит АБ в снижении вероятности рецидива заболевания.
Разрешение симптомов сопровождается пополнением кластеров Bacteroidetes и Clostridium IV и XIVa с соответствующим снижением Proteobacteria. Недавнее исследование детей с Clostridium-ассоциированной диареей продемонстрировало 94, 75 и 54% успешного выздоровления после в/кишечной трансплантации фекалий из банка донорского стула у ранее здоровых детей, детей со сложными медицинскими показаниями и детей с ВЗК соответственно.
6. Микробном и связь с воспалительным заболеванием кишечника. Болезнь Крона и язвенный колит — хронические ВЗК, результат сочетания измененной восприимчивости хозяина и дисбактериоза, т.е. изменения микробиоты кишечника. Исследования показывают, что уровень конкордантности у монозиготных близнецов составляет 10-15% при язвенном колите и 30-35% при болезни Крона. Это демонстрирует генетический компонент каждого заболевания и выделяет факторы окружающей среды, которые вызывают и стимулируют прогрессирование заболевания. Более 150 однонуклеотидных полиморфизмов (SNP; англ. Single Nucleotide Polymorphism) связаны с заболеваниями, выявляя потенциальные дефекты в геноме микроорганизмов. Некоторые микроорганизмы участвуют в поддержании барьерной функции, в работе врожденного иммунитета, аутофагии, адаптивном иммунитете, а также в метаболизме и клеточном гомеостазе.
При ВЗК микробиота изменяется во всем кишечном тракте. Несмотря на описанную значительную гетерогенность, ВЗК часто сопровождается уменьшением количества бактероидов, клостридий, бифидобактерий и фирмикутов. Описаны также разрастания Е. coli и других Enterobacteriaceae. Увеличение количества метаболизаторов серы характерно ВЗК. АБ и биологическая терапия (моноклональные АТл, направленные на нейтрализацию ФНО) использовались для лечения дисбактериоза и воспалительной реакции при ВЗК. Проводятся испытания трансплантации фекалий, чтобы определить, может ли невоспалительная микробиота от здорового донора смягчить симптомы и прогрессирование ВЗК.
7. Микробиом и ожирение. Ожирение и метаболический синдром связаны с заметными изменениями в микробиоме кишечника, касающимися состава и метаболической функции, что приводит к большему извлечению энергии из рациона. Раннее исследование микробиома при ожирении зафиксировало увеличение соотношения между типами Firmicutes: Bacteroidetes, но дискуссии по поводу специфических для ожирения изменений в микробиоме продолжаются. Многочисленные исследования продемонстрировали снижение соотношения Firmicutes: Bacteroidetes в фекальной микробиоте у тучных людей по сравнению с контрольной группой нормального телосложения. Дальнейшие исследования показывают, что пропорции групп на уровне биологического типа менее важны, чем изменения в подгруппах Firmicutes, которые продуцируют бутират.
Микробиом кишечника усиливает извлечение калорий из неперевариваемых субстратов рациона (например, полисахариды). Микробиом производит разлагающие ферменты для расщепления этих субстратов: некоторые гликозилгидролазы не кодируются в геноме человека. Молекулярные исследования показывают, что микробиом кишечника может взаимодействовать с эпителием кишечника, изменяя общий энергетический гомеостаз и накопление жира. Например, микробиом кишечника может продуцировать SCFAs, которые изменяют экспрессию эндокринных пептидов (глюкагоноподобный пептид-1 и пептид YY), влияющих на гомеостаз глюкозы и насыщение. За счет производства SCFA и кетонов микробиота может изменять симпатический тонус. Определенные микробиомы подавляют другие, индуцируя жировой фактор (также называемый ангиопоэтин-подобным белком 4), ингибитор липопротеинипазы кишечного, печеночного и жирового происхождения.
Колонизация разнообразной микробиотой подавляет экспрессию жирового фактора, и диетические добавки с Lactobacillus paracaseifur подавляют уровни индуцированной экспрессии жирового фактора. У мышей, получавших западную диету, развилось ожирение, которое передавалось худым мышам-реципиентам после трансплантации микробиоты мышей с ожирением. И наоборот, у мышей с ожирением, получавших АБ, наблюдалась меньшая инсулинорезистентность, более низкие гликемические индексы натощак и улучшенная толерантность к глюкозе по сравнению с др. группой, что указывает на участие микробиома в этих физиологических изменениях.
8. Микробиом при недоедании. Недоедание — основная причина заболеваемости и смертности во всем мире. В тяжелой форме недоедание может привести к квашиоркору, который характеризуется генерализованным отеком, анорексией, ожирением печени, изъязвлениями кожи и раздражительностью. Готовые к употреблению продукты распределяются для восстановления питания в районах с жесткими ограничениями в питании.
Моно- и дизиготные близнецы в Малави изучались на предмет изменений в микробиоме, связанных с умеренным и тяжелым недоеданием, включая квашиоркор. Среди близнецов с разной степенью недоедания у близнецов с ранее существовавшим легким недоеданием кишечная микробиота значительно изменилась в процессе приема добавок. Напротив, у близнецов с ранее существовавшим квашиоркором микробиота слабо или вообще не реагировала на пищевые добавки. Эти результаты были обобщены после трансплантации микробиоты близнецов ранее стерильным мышам. Мыши, получившие микробиоту малавийских близнецов с квашиоркором, испытали более резкую потерю веса на диете малавийского типа и более быструю потерю веса после приема готовых пищевых добавок, чем мыши, которым пересаживали фекалии здоровых близнецов.
У мышей с трансплантированной микробиотой квашиоркора наблюдались проблемы с метаболизмом углеводов, липидов и аминокислот, несмотря на пищевые добавки малавийской диеты. Эти данные указывают на то, что тяжелое недоедание — результат сочетания дефицита питания и микробиома с измененными метаболическими возможностями, которые сложно восстановить с помощью лечебных добавок.
д) Терапевтическое воздействие на микробиом. Терапевтическое воздействие на микробиом делится на 6 основных категорий: АБ, пребиотики, пробиотики, синбиотики, постбиотики и трансплантация фекалий (см. Clostridium-ассоциированная диарея и ВЗК). Постбиотики — нежизнеспособные микробные компоненты или метаболиты, которые могут изменять микробиоту или вызывать физиологические изменения в организме хозяина. На текущий момент недостаточно данных для обсуждения постбиотической терапии.
1. Пребиотики. Пребиотики — неперевариваемые пищевые компоненты, которые благотворно влияют на организм, избирательно стимулируя рост или активность одного или ограниченного числа бактерий в толстой кишке. АБТ истощают часть микробиоты, пребиотики способствовуют росту полезных организмов (бифидо- и лактобактерии). Пребиотики — углеводы, олигосахариды, которые могут избирательно метаболизироваться составляющими микробиоты. Они стимулируют рост желательных организмов и могут катаболизироваться с образованием полезных конечных продуктов (SCFA), которые используются в качестве энергетических субстратов эпителием кишечника. Естественные пребиотические олигосахариды содержатся в грудном молоке и используются в качестве добавок к грудному молоку и смеси. Введение пребиотиков доношенным детям продемонстрировало ожидаемый рост бактерий, но клинически значимые преимущества от приема пребиотиков четко не установлены.
Лечение доношенных детей фруктоолигосахаридами увеличивает количество фекальных бифидобактерий, но не влияет на рост ребенка, несмотря на повышение содержания SCFAs в фекальной массе. Систематический обзор темы дал аналогичный вывод. У недоношенных детей отмечается снижение уровня или полное отсутствие бифидобактерий и лактобактерий в кишечном тракте, несмотря на полноценное грудное молоко. Для увеличения бактериальных популяции в кишечном тракте недоношенных детей применяются пребиотики. Предполагаемое преимущество — снижение риска развития некротического энетроколита. Рандомизированные исследования с надлежащей популяцией для проверки этой гипотезы не проводились.
2. Пробиотики. Пробиотики — изоляты микробиоты человека (жизнеспособные микроорганизмы). В качестве альтернативы можно принимать пробиотики для повышения уровня содержания микроорганизма, уже присутствующего в микробиоте. Пробиотики вводят перорально или в виде вагинальных суппозиториев. Многие роды и виды бактерий и грибов были изучены на предмет пробиотических эффектов. Общие бактериальные роды — бифидобактерии, лактобациллы, стрептококки, энтерококки и кишечная палочка. Меньшее количество небактериальных организмов изучено на предмет пробиотических эффектов. У Saccharomyces boulardii, относящихся к пекарским дрожжам (Saccharomyces cerevisiae), отмечены определенные положительные пробиотические эффекты. Эти пробиотические микроорганизмы не следует путать с более патогенными штаммами в пределах их родов и видов.
Бифидобактерии и лактобациллы часто встречаются в грудном молоке и кале у младенцев с низкой частотой диарейных заболеваний и аллергии. За исключением людей со значительным иммунодефицитом, серьезно нарушенными барьерами слизистой оболочки или ЦВК (многие микроорганизмы могут абсорбироваться на пластике), бактериальные пробиотики оказались относительно безопасными даже при наличии млрд КОЕ. Частые побочные эффекты, связанные с пробиотиками, — спазмы в животе, тошнота, лихорадка, изменения стула, метеоризм и нарушение вкуса. Хотя бактериальные пробиотики широко применялись среди людей, доказательства их эффективности ограничены небольшим числом состояний. Пробиотики демонстрируют эффективность при определенных состояниях, включая диарею, вызванную АБ, при профилактике и снижении атопии у детей из группы высокого риска, а также сокращении продолжительности и рецидивов инфекции С. difficile. Испытания показывают снижение частоты некротического энтероколита среди недоношенных детей.
Пробиотики могут снизить риск респираторных инфекций и рецидивов ИМП, одновременно уменьшая симптомы и частоту обострений ВЗК. Уменьшается частота и продолжительность диареи, связанной с приемом АБ. Метаанализ показал RR диареи, связанной с АБ, при введении пробиотика 0,58 (95% доверительный интервал 0,05-0,68) среди комбинированных исследований с использованием Lactobacillus, Bifidobacterium, Saccharomyces, Streptococcus, Enterococcus и/или Bacillus. Введение комбинаций организмов не приводит к большей эффективности.
Метаанализ, посвященный эффективности пробиотиков в снижении заболеваемости Clostridium-ассоциированной диареи, продемонстрировал умеренные доказательства для этой практики. Согласно анализу, в >1800 исследований, в т. ч. из педиатрической популяции, пробиотики снижали риск Clostridium-ассоциированной диареи на 64% с RR 0,36 (95% доверительный интервал 0,26-0,51). Педиатрическая подгруппа была проанализирована в соответствующих исследованиях, выявив пользу у педиатрических пациентов и здоровых детей (RR 0,37; 95% доверительный интервал 0,23-0,60). Был использован ряд пробиотиков, включая различные штаммы Lactobacillus и S. boulardii.
Проведено >15 испытаний для изучения влияния пробиотиков на организм плода во время беременности и у младенцев на профилактику атопического дерматита. Метаанализ предполагает умеренную пользу от введения пробиотиков для предотвращения атопического дерматита. Исследования включали введение Lactobacillus rhamnosus только беременной или младенцу, либо обоим. Общий RR 0,79 (95% доверительный интервал 0,71-0,88) был постоянным независимо от лечения матери, ребенка или обоих. Продолжительность составляла >6 мес, но длительность терапии незначительно изменяла эффект. RR аналогичен для профилактики IgE- и He-IgE-ассоциированного атопического дерматита. Симбиотики — комбинации пробиотика и пребиотика. Большое двойное слепое плацебо-контролируемое исследование с участием >4500 младенцев в Индии показало, что ежедневный прием перорального симбиотика Lactobacillus plantarum и фруктоолигосахарида в неонатальном периоде приводит к значительному снижению сепсиса, пневмонии, кожных инфекций и общей смертности.