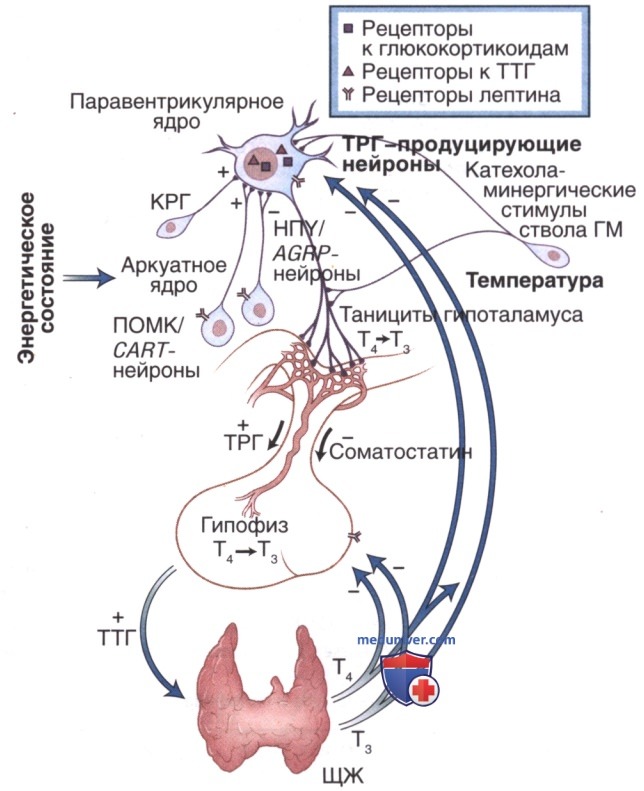
Гипофиз является основным регулятором сложной гормональной системы. Гипофиз получает сигналы от гипоталамуса и в ответ синтезирует гормоны, влияющие на периферические эндокринные железы. Периферические эндокринные железы производят гормоны, которые по принципу отрицательной обратной связи подавляют секрецию на уровне гипоталамуса и гипофиза (рис. 1, 2).
Механизм обратной связи позволяет гипофизу регулировать количество гормона, высвобождаемого в кровеносное русло периферическими эндокринными железами. Центральная роль гипофиза в гормональной системе и его способность интерпретировать и отвечать на различные сигналы привели к тому, что в англоязычной литературе к нему стали применять термин «master gland». В табл. 1 представлен список гормонов гипоталамуса и гипофиза и их функции.
а) Анатомия. Гипофиз располагается в основании черепа в седловидной ямке клиновидной кости, называемой турецким седлом. Костная структура защищает и окружает гипофиз с двух сторон и снизу. Твердая оболочка, слой из плотной соединительной ткани, формирует крышу седла. Наружный листок твердой оболочки продолжается в седло и выстилает его поверхность. Таким образом, гипофиз расположен экстрадурально и обычно не контактирует со СМЖ.
Питуитарная железа соединена с гипоталамусом воронкой гипофиза. Гипофиз состоит из передней (аденогипофиз) и задней (нейрогипофиз) долей. Передняя доля составляет 80% процентов всей железы.
б) Эмбриология. Передняя доля гипофиза происходит из выпячивания эктодермы ротовой полости — кармана Ратке. Он отделяется от ротового эпителия и становится самостоятельной структурой с быстро пролиферирующими клетками. К 6-й неделе гестации карман Ратке полностью утрачивает связь с глоткой и напрямую связывается с распространяющимся книзу гипоталамусом, что приводит к формированию ножки гипофиза.
Из остатков ткани между карманом Ратке и ротовой полостью могут развиваться краниофарингиомы — самые частые опухоли этой области.
в) Кровоснабжение. Артериальное кровоснабжение гипофиза происходит из внутренней сонной артерии через нижнюю, среднюю и верхнюю гипофизарную артерии. Сеть сосудов формирует уникальную портальную систему кровообращения, связывающую гипоталамус и гипофиз. Ветви верхних гипофизарных артерий проникают в воронку и формируют сеть сосудов, которая проходит через воронку гипофиза и заканчивается сетью капилляров в передней доле.
С помощью этой портальной венозной системы гормоны гипоталамуса доставляются в переднюю долю гипофиза. Гормоны передней доли гипофиза, в свою очередь, секретируются во вторичные портальные вены, которые впадают в венозные синусы ТМО.
г) Типы клеток передней доли гипофиза. Дифференцировка и пролиферация разных типов клеток передней доли гипофиза осуществляется под действием ряда последовательно синтезируемых белковых факторов транскрипции. Эти белки являются членами большого семейства ДНК-связывающих белков, напоминающих белки, кодируемые гомеобокс-генами.
Последствия мутаций некоторых из этих генов проявляются у человека множественной недостаточностью гормонов гипофиза. Пять типов клеток передней доли гипофиза продуцируют шесть пептидных гормонов. Соматотрофы продуцируют гормон роста (ГР), лактотропы — пролактин, тиреотропы синтезируют ТТГ, кортикотропы — проопиомеланокортин (ПОМК), предшественник АКТГ, а гонадотропы производят ЛГ и ФСГ.
1. Гормон роста. Человеческий ГР представляет собой одноцепочечный полипептид, состоящий из 191 аминокислоты, который синтезируется, хранится и секретируется соматотрофами в гипофизе. Его ген (GH1) — первый в кластере пяти близкородственных генов, которые находятся на длинном плече 17-й хромосомы (q22-24). Остальные четыре гена (CS1, CS2, GH2 и CSP) имеют >90% последовательностей, идентичных с геном GH1.
ГР поступает в кровеносное русло импульсно под контролем гормонов гипоталамуса. Периодический характер секреции ГР-рилизинг-гормона, стимулирующего высвобождение ГР, и соматостатина, ингибирующего секрецию ГР, приводит к ритмической секреции ГР. Пики секреции ГР возникают, когда пики ГР-рилизинг-гормона совпадают с минимальным выбросом соматостатина. Грелин, белок, который производится в дугообразном ядре гипоталамуса и в значительно большем количестве в желудке, также стимулирует выброс ГР.
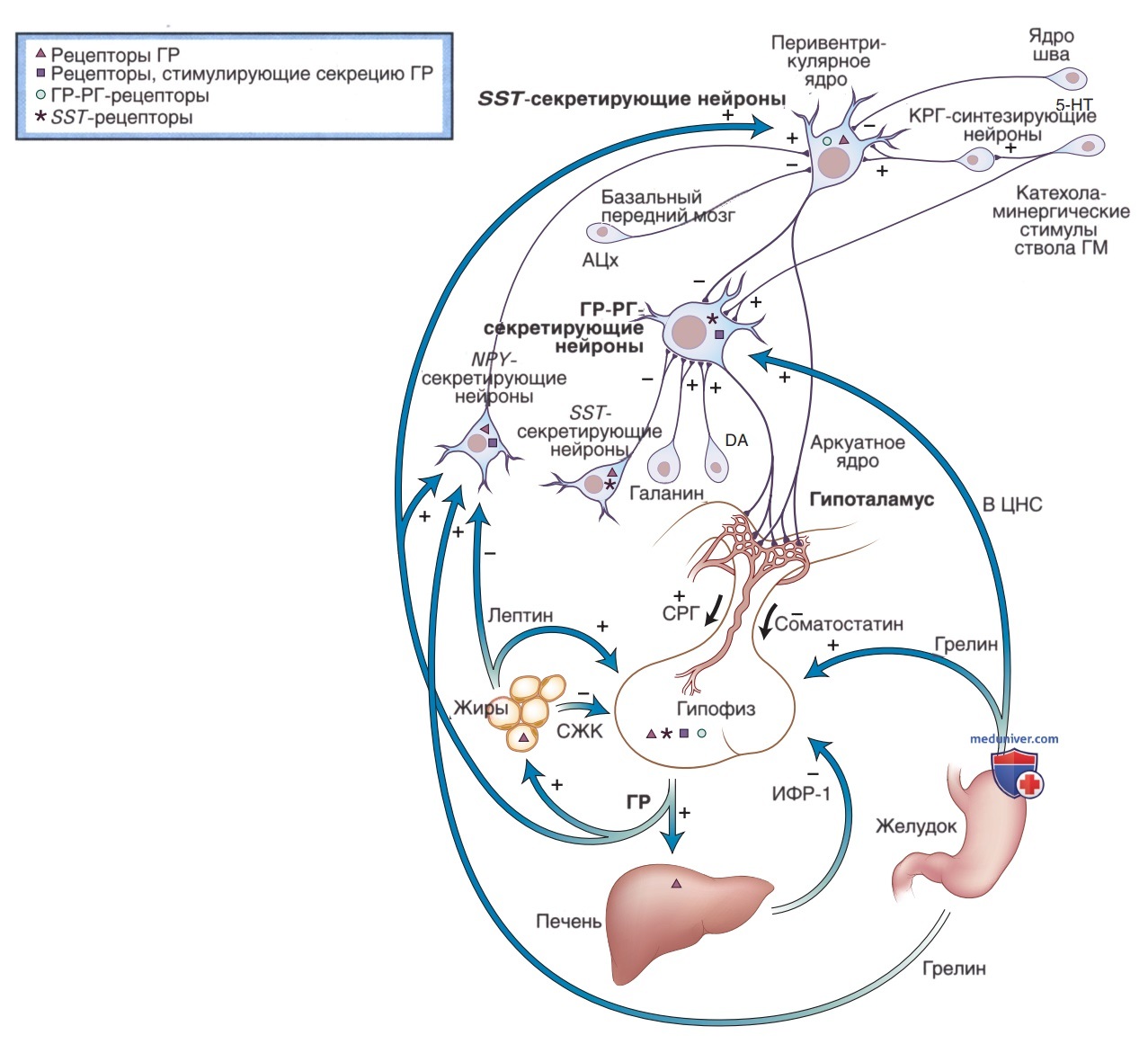
Помимо трех гипоталамических гормонов в стимуляции и подавлении секреции гормона роста играют роль и физиологические факторы. Сон, упражнения, физический стресс, травма, острое заболевание, половое созревание, голодание и гипогликемия стимулируют выработку ГР, в то время как гипергликемия, гипотиреоз и глюкокортикоиды ингибируют выработку ГР (рис. 3).
Рисунок 3. Регуляция оси гипоталамус-гипофиз-гормон роста (ГР). Секреция ГР гипофизом стимулируется ГР-рилизинг-гормоном (ГР-РГ) и ингибируется соматостатином. Инсулиноподобный фактор роста (ИФР-1) и свободные жирные кислоты по системе отрицательной обратной связи осуществляют контроль над секрецией ГР в гипоталамусе. Сам ГР по короткой петле отрицательной обратной связи активирует нейроны, секретирующие соматостатин, в перивентрикулярном ядре гипоталамуса. Эти нейроны, секретирующие соматостатин, непосредственно контактируют с нейронами дугообразного ядра, секретирующими ГР-РГ, а коллатерали аксонов идут к медиальному возвышению. Нейроны, секретирующие нейропептид Y (НПY) в дугообразном ядре, также опосредованно снижают частоту выделения ГР, интегрируя периферические сигналы ГР, лептина и грелина и передавая их в перивентрикулярные нейроны, секретирующие соматостатин. Грелин выделяется в желудке и является естественным лигандом рецептора, стимулирующего секрецию ГР (GHS), который увеличивает выработку ГР как на гипоталамическом, так и на гипофизарном уровнях. На основании косвенных фармакологических данных установлено, что высвобождение ГР-РГ стимулируется галанином, γ-аминомасляной кислотой, α2-адренергическими и дофаминергическими стимулами и ингибируется соматостатином. Лиганды мускаринового ацетилхолинового и 5-HT-1D рецепторов ингибируют выработку соматостатина, а β2-адренергические стимулы и кортикотропин-рилизинг-гормон стимулируют. ЦНС — центральная нервная система; DA — дофамин; 5-НТ — серотонин (5-гидрокситриптамин)
ГР связывается с молекулами рецепторов на поверхности клетки-мишени. Рецептор ГР представляет собой одноцепочечную молекулу из 620 аминокислот с тремя доменами: внеклеточным, одним трансмембранным и цитоплазматическим. Протеолитически расщепленные фрагменты внеклеточного домена циркулируют в плазме и играют роль ГР-связывающего белка. Как и другие члены семейства цитокиновых рецепторов, цитоплазматический домен рецептора ГР лишен внутренней киназной активности; вместо этого при связывании ГР происходит димеризация рецептора и активация рецептор-ассоциированной киназы Janus (Jak2).
Фосфорилирование киназы и других белковых субстратов запускает последовательность реакций, которые приводят к изменениям транскрипции ядерных генов. Трансдуктор сигнала и активатор транскрипции 5b (STAT5b) играет ключевую роль во взаимосвязи между активацией рецепторов и изменениями транскрипции генов.
Биологические эффекты ГР включают в себя ускорение линейного роста, утолщение костей, рост мягких тканей, синтез белка, высвобождение липидов из жировой ткани, повышение инсулинорезистентности и уровня глюкозы крови. Митогенное действие ГР опосредовано увеличением синтеза инсулиноподобного фактора роста 1 (ИФР-1), прежде носившего название соматомедин С, одноцепочечного пептида, состоящего из 70 аминокислот, который кодирует ген, расположенный на длинном плече 12-й хромосомы.
ИФР-1 имеет значительную гомологию с инсулином. Циркулирующий ИФР-1 синтезируется в основном в печени. Кроме того, он локально образуется в мезодермальных и эктодермальных клетках, особенно в ростовых зонах костей у детей, где действует посредством паракринных и аутокринных механизмов. Уровень циркулирующего в крови ИФР-1 связан с уровнем ГР и нутритивным статусом. В крови ИФР-1 связывается с ИФР-связывающими белками. Основным из них является комплекс с молекулярным весом 150 кДа (ИФРСБ-3), уровень которого снижен у детей с дефицитом ГР.
Человеческий рекомбинантный ИФР-1 может найти применение при периферической резистентности к ГР, напр., при синдроме Ларона, а также при образовании АТл к экзогенному ГР. ИФР-2 — одноцепочечный белок, состоит из 67 аминокислот, его кодирует ген, расположенный на коротком плече 11-й хромосомы. У него есть гомология с ИФР-1. О его физиологической роли известно гораздо меньше, однако, по всей видимости, он является важным митогеном в клетках костной ткани, где его концентрация во много раз превышает уровни ИФР-1.
2. Пролактин. Пролактин — белок, состоящий из 199 аминокислот, продуцируемый лактотрофами гипофиза. Регуляция выработки пролактина уникальна потому, что он секретируется постоянно, пока его синтез не ингибируется дофамином, белком, который вырабатывают нейроны гипоталамуса. Повреждение гипоталамуса или воронки гипофиза может привести к повышению уровня пролактина. Антагонисты дофамина, первичный гипотиреоз, прием ТРГ и ЗНО гипофиза могут приводить к повышению уровня пролактина в плазме крови. Агонисты дофамина и повреждение гипофиза являются причинами снижения уровня пролактина.
Основная физиологическая роль пролактина — инициация и поддержание лактации. Пролактин подготавливает МЖ к лактации и стимулирует образование молока в послеродовый период. Во время беременности пролактин стимулирует развитие грудных протоков, однако лактация не наступает из-за высоких уровней эстрогена и прогестерона. После рождения ребенка уровни эстрогена и прогестерона падают, а физиологические стимулы, такие как сосание и стимуляция соска, поддерживают выработку пролактина и запускают лактацию.
3. Тиреотропный гормон. ТТГ состоит из двух гликопротеидных цепей (α, β), связанных водородной связью: α-субъединица состоит из 89 аминокислот и идентична таковой в других гликопротеидных гормонах (ФСГ, ЛГ и ХГЧ), а β-субъединица, состоящая из 112 аминокислот, специфична для ТТГ.
ТТГ хранится в секреторных гранулах и высвобождается в сосудистое русло главным образом в ответ на ТРГ, который вырабатывается в гипоталамусе. ТРГ секретируется гипоталамусом в гипоталамо-гипофизарную портальную систему и стимулирует высвобождение ТТГ из тиреотрофов гипофиза. ТТГ стимулирует секрецию Т4 и Т3 ЩЖ через образование циклического аденозинмонофосфата и активации системы вторичных мессенджеров, связанных с G-белком. Ингибирование по принципу обратной связи в основном осуществляется Т3, кроме того, секрецию ТРГ и ТТГ тормозят дофамин, соматостатин и глюкокортикоиды.
Дефицит ТТГ приводит к потере функции и атрофии ЩЖ, в то время как при избытке ТТГ наблюдается ее гипертрофия и гиперплазия.
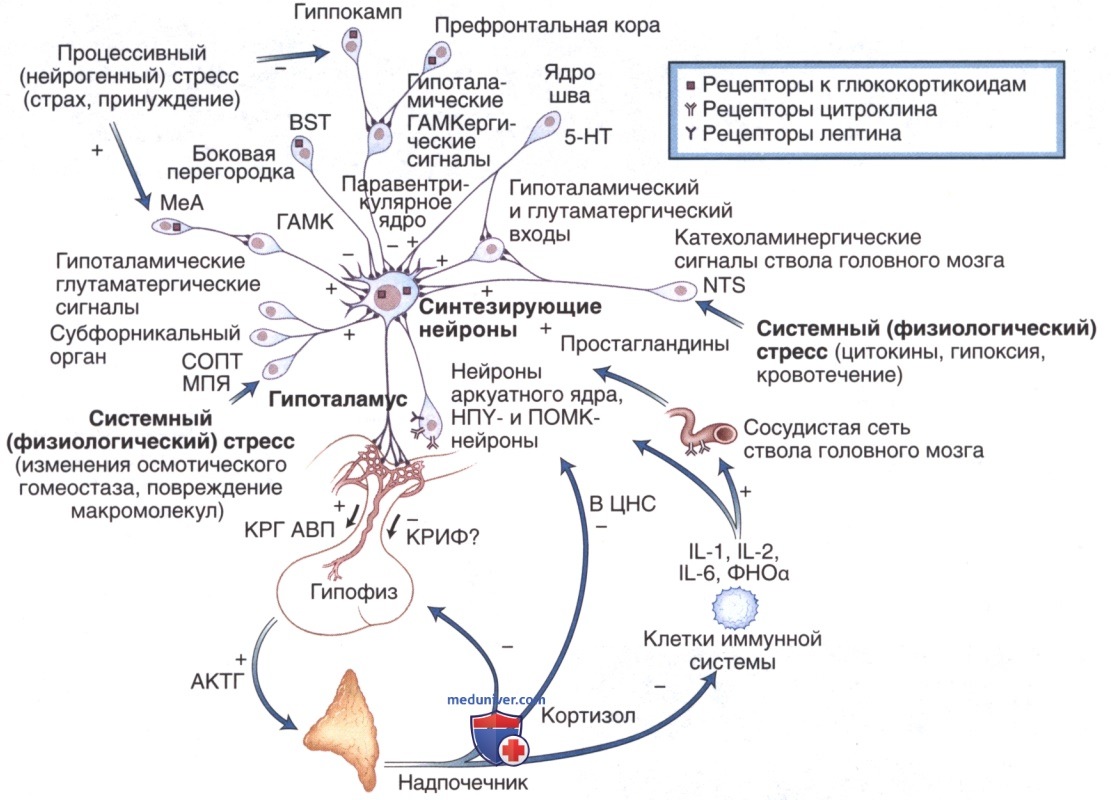
4. Адренокортикотропный гормон. АКТГ — одноцепочечный пептид, состоящий из 39 аминокислот, образуется при протеолитическом расщеплении ПОМК. ПОМК синтезируется в гипофизе, состоит из 240 аминокислот и является гликопротеидным предшественником. Кроме АКТГ, ПОМК также содержит последовательности для образования липотропинов, меланоцитстимулирующих гормонов (α, β, γ) и β-эндорфина.
Выработка АКТГ регулируется кортикотропин-рилизинг-гормоном (КРГ), пептидом из 41 аминокислоты, который преимущественно обнаруживается в области срединного возвышения, но присутствует и в других отделах ГМ, а также за его пределами. Для АКТГ характерен суточный ритм секреции. Он влияет на кору надпочечников и стимулирует синтез и секрецию кортизола. Самые высокие уровни АКТГ и кортизола определяются утром в момент пробуждения, затем снижаются к концу дня, достигая самого низкого уровня через 1-2 ч после начала сна. У человека, судя по всему, АКТГ играет основную роль в стимуляции пигментации.
Подобно ТРГ и ТТГ, КРГ и АКТГ реализуют свое действие посредством образования циклического аденозинмонофосфата и активируют систему вторичных мессенджеров, связанных с G-белком. Хотя КРГ является основным регулятором секреции АКТГ, другие гормоны также могут влиять на его выработку. Аргинин-вазопрессин, окситоцин, ангиотензин II и холецистокинин стимулируют выработку КРГ и АКТГ, в то время как предсердный натрийуретический пептид и опиоиды оказывают ингибирующее действие. По аналогии с обратной связью между Т3 и ТТГ и ТРГ, кортизол тормозит выработку КРГ и АКТГ. Физиологические состояния, такие как стресс, голодание и гипогликемии, также стимулируют выработку КРГ и АКТГ.
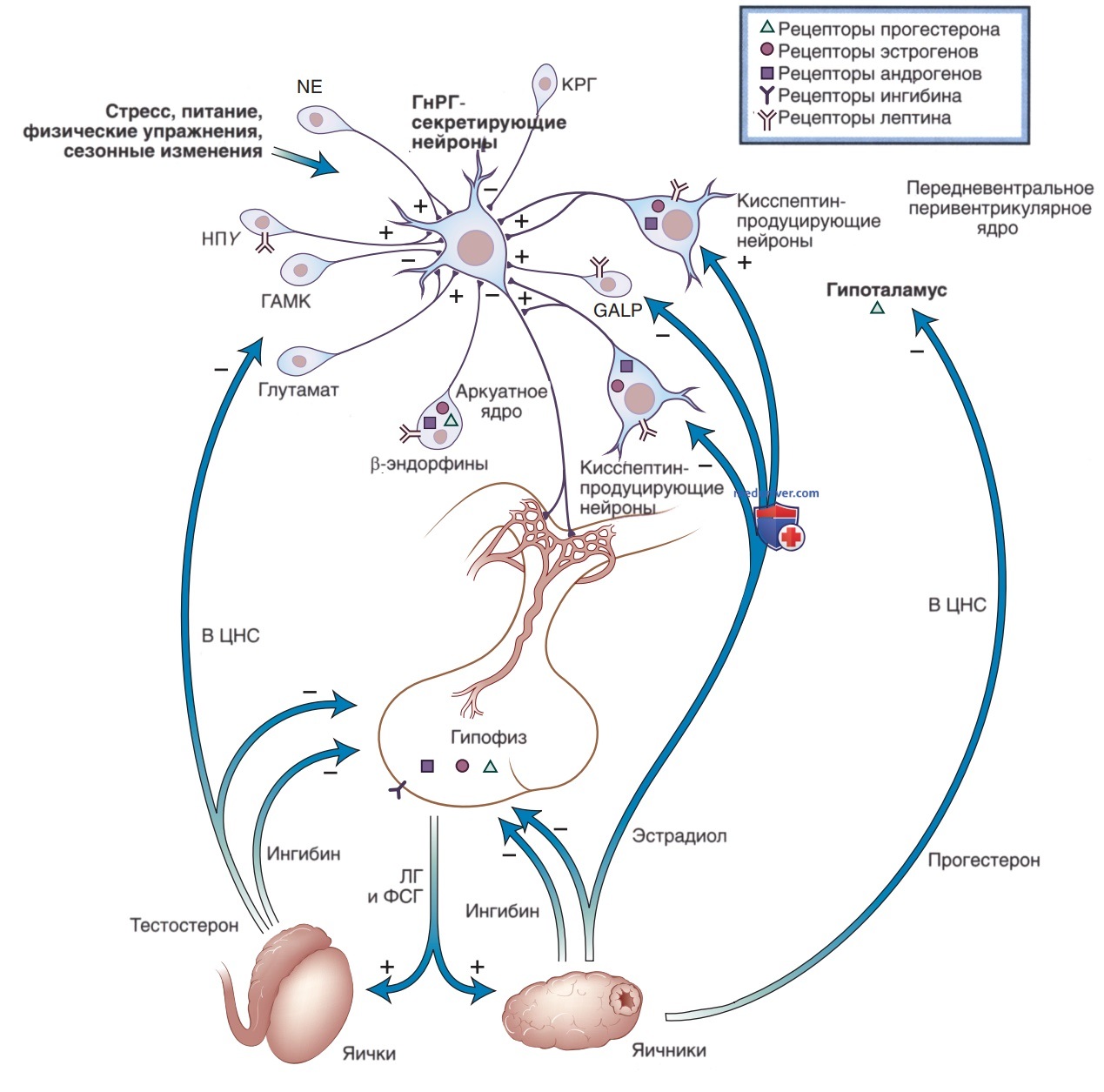
5. Лютеинизирующий гормон и фолликулостимулирующий гормон. Гонадотропные гормоны включают в себя два гликопротеида — ЛГ и ФСГ. Они содержат ту же α-субъединицу, что и ТТГ и ХГЧ, но разные β-субъединицы. Рецепторы ФСГ на клетках гранулезы яичников и клетках Сертоли яичек активируют ФСГ-стимулированные процессы фолликулогенеза и гаметогенеза. Связываясь со специфическими рецепторами на поверхности клеток теки яичника и клеток Лейдига яичек, ЛГ способствует образованию желтого тела и функционированию клеток Лейдига (рис. 4). Рецепторы ЛГ и ФСГ принадлежат к классу рецепторов с семью трансмембранными доменами.
Рисунок 4. Регуляция гипоталамо-гипофизарно-гонадной оси. Схематическая иллюстрация гипоталамо-гипофизарно-гонадной оси, демонстрирующая нейрональные системы, которые регулируют секрецию гонадотропин-рилизинг-гормона (ГнРГ) и обратную связь гонадных стероидов на уровне гипоталамуса и гипофиза. ЦНС — центральная нервная система; КРГ — кортикотропин-рилизинг-гормон; ФСГ — фолликулостимулирующий гормон; ГАМК — γ-аминомасляная кислота; GALP — галанин-подобный пептид; ЛГ — лютеинизирующий гормон; NE — норэпинефрин; НПY — нейропептид Y
Связанный с лигандом (гонадотропными гормонами) рецептор, взаимодействуя с G-белком, активирует его, в результате происходит связывание последнего с аденилатциклазой и ее активация.
ЛГ-рилизинг-гормон представляет собой декапептид. Он был выделен, синтезирован и на данный момент широко используется в клинических исследованиях. Поскольку он приводит к выработке ЛГ и ФСГ в одних и тех же клетках-гонадотрофах, судя по всему, он является единственным гонадотропин-рилизинг-гормоном.
Секреция ЛГ подавляется андрогенами и эстрогенами, а секреция ФСГ — продукцией ингибина гонадами. Ингибин — гликопротеид с молекулярной массой 31 кДа, который вырабатывается клетками Сертоли. Ингибин состоит из α- и β-субъединиц, связанных дисульфидными мостиками. Существует также β-β-димер (активин), однако его биологический эффект заключается в стимуляции секреции ФСГ. В настоящее время описаны биологические особенности этих недавно идентифицированных гормонов.
Помимо эндокринного влияния активин оказывает паракринное действие в яичках. Он способствует ЛГ-индуцированной выработке тестостерона, что указывает на прямое влияние клеток Сертоли на клетки Лейдига.
д) Клетки задней доли гипофиза. Задняя доля гипофиза является частью функциональной единицы — нейрогипофиза, который состоит из нейронов супраоптического и паравентрикулярного ядер гипоталамуса; аксонов нейронов, формирующих воронку гипофиза; а также терминалей нейронов в срединном возвышении или задней доле. Аргинин-вазопрессин (АДГ) и окситоцин — это два гормона, образующиеся в результате нейросекреции гипоталамических ядер. Они оба высвобождаются из задней доли гипофиза и представляют собой октапептиды, отличающиеся всего двумя аминокислотами.
1. Антидиуретический гормон. АДГ отвечает за сохранение жидкости в организме на уровне почек, повышая проницаемость собирательных трубочек почек для воды. АДГ стимулирует транслокацию водных каналов посредством своего взаимодействия с рецепторами вазопрессина-2 в собирательном протоке, которые действуют через G-белки, повышая активность аденилилциклазы и проницаемость для воды. V2-рецепторы также опосредованно влияют на активность фактора фон Виллебранда и тканевого активатора плазминогена. В высоких концентрациях АДГ активирует V1-рецепторы в гладкомышечных клетках и гепатоцитах, оказывая сосудосуживающее и гликогенолитическое действие путем мобилизации в/клеточных запасов кальция.
Отдельные V3-рецепторы способствуют повышению секреции АКТГ. Эти эффекты в большей степени запускают гидролиз фосфатидилинозитола, а не продукцию циклического аденозинмонофосфата.
АДГ и сопутствующий ему белок нейрофизин II кодируются одним и тем же геном. В результате расщепления общего препрогормона образуются два гормона, которые транспортируются в нейросекреторных гранулах в заднюю долю гипофиза. Оба гормона секретируются в эквимолярном количестве.
АДГ имеет короткий период полураспада и быстро реагирует на изменение водного баланса организма. Его секреция стимулируется повышением осмоляльности плазмы, воспринимаемой осморецепторами гипоталамуса, и снижением ОЦК, воспринимаемым барорецепторами каротидного синуса дуги аорты.
2. Окситоцин. Окситоцин стимулирует сокращения матки в ответ на растяжение родовых путей во время прелиминарного периода и родов, а также сокращения гладкомышечной мускулатуры груди во время сосания, что приводит к выделению молока. Исследования показывают, что окситоцин также играет определенную роль в оргазме, социальном признании, связывании пар, тревоге, доверии, любви и материнском поведении. Не так давно стало известно, что посредством взаимодействия с G-белок-связанными рецепторами в ПЖЖ и жировой ткани окситоцин играет значительную роль в регуляции аппетита и ожирения, вызывая анорексию.