Анатомия и физиология вентрального кохлеарного ядра (ВКЯ)
Как описано отдельной статье «Передача звука во внутреннем ухе в норме», сложный звук распадается на составные частотные компоненты путем механической настройкой улитки. Поскольку акустическая энергия тонотопически распределена вдоль кохлеарной части, выходные частоты отдельных волокон слухового нерва зависят от формы волны. Дополнительный процесс восстановления звукового сигнала из рассеянных периферических сигналов начинается в улитковом ядре, где параллельные потоки восходящей информации, каждый с собственной уникальной функциональной ролью, представлены сходящимися структурами волокон слухового нерва.
Следующее обсуждение строится вокруг самой широкой функциональной дихотомной структуры: вентрального и дорсального разделения комплекса улитковых ядер. Это разделение возникло в самом начале анатомического формирования слухового отдела ствола мозга. Такое разделение сохранилось лишь по причине того, что представленная клеточная морфология обеспечила превосходное функционирование на начальных этапах центральной слуховой обработки.
Вентральное кохлеарное ядро (ВКЯ) служит первичной точкой входа для эфферентных путей к верхней оливе, латеральной петле и нижнему коленцу (НК). Слуховая информация идет к каждой из этих структур в определенных проводящих путях, которые приспособлены для селективного кодирования фаз и уровней частотных компонентов, а также и более глобальных свойств, таких как амплитуда модуляции стимула. Это функциональное разделение происходит из-за различий в синаптических структурах, конвергирующих структурах и внутренних электрических свойств нейронов ВКЯ.
а) Анатомия вентрального кохлеарного ядра. Приблизительно 30000 слуховых нервных волокон соединяют человеческое внутреннее ухо с комплексом кохлеарных ядер. После поступления в ствол мозга пути разделяются на восходящую ветвь, которая идет к переднему ВКЯ и нисходящую ветвь, которая проходит через заднее ВКЯ и дорсальное кохлеарное ядро (ДКЯ). Хотя расположение проекций повторяет тонотопическую организацию улиткового разделения, линейное отображение частоты преобразуется в конечные поля. Посредством каждой двухмерной частотной пластинки ортогональная ось кодирует другие показатели звука.
Завершение слухового нерва в кохлеарном ядре имеет принципиальное значение. Вся информация о звуке должна быть передана к вышестоящим центрам посредством выходных частот слухового ядра. Для каждого слухового нервного волокна есть примерно два нейрона ВКЯ, что предполагает высокую степень схождения в ядре.
Каким образом нейроны будут интегрировать многообразные входящие сигналы слухового нерва, зависит от физических характеристик синапсов улитковых ядер. Известны две базовые структуры. Нейроны с широкими интеграционными свойствами имеют обширные дендритные поля, покрытые шишковидными окончаниями большого числа волокон слухового нерва. Нейроны, которые точно сохраняют образцы сигналов слухового нерва, управляются большими аксональными окончаниями, концевыми утолщениями Хелда. Эти входы малочисленны, но по отдельности достаточно мощные, чтобы активизировать постсинаптическую активность.
Основные подтипы нейронов ВКЯ отличаются тремя основными типами клеток. Мшистые клетки имеют мелкие, похожие на куст, дендриты, глобулярные или сферические тела клеток и синаптические концевые утолщения. Как большинство нейронов в центральной нервной системе, мультиполярные (или звездчатые), клетки имеют длинные, практически без ветвей, дендриты и шишковидные синапсы. Интеграционные свойства этих клеток прежде всего определены тем, лежат ли их дендритные области в пределах плоскости частотной пластины или проходят через нее. Так называемые «осьминогообразные» клетки имеют длинные дендриты, с пучками от одной стороны тела клетки, напоминая головоногих.
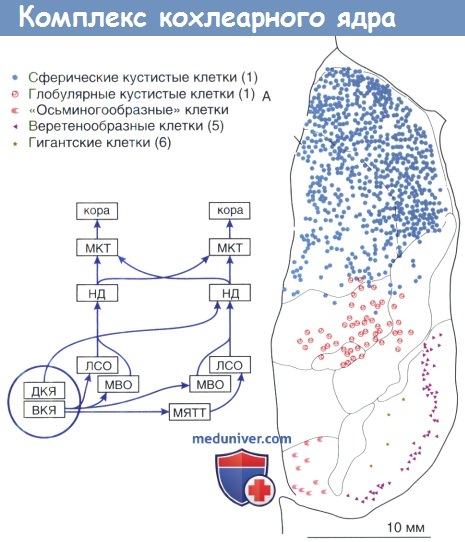
Цитоархитектоника ВКЯ организована регионально. Сферические мшистые клетки, располагаясь в самом переднем отделе, окружают точку входа слухового нерва. Глобулярные мшистые клетки занимают промежуточное местоположение, в то время как «осьминогообразные» клетки расположены в заднем отделе. Мультиполярные клетки, как наиболее структурно разнообразные нейроны, в основном рассеяны.
Топографическая группировка морфологических нейрональных классов в комплексе кохлеарного ядра.
Схема показывает относительное положение кохлеарного ядра в пределах центральных слуховых путей.
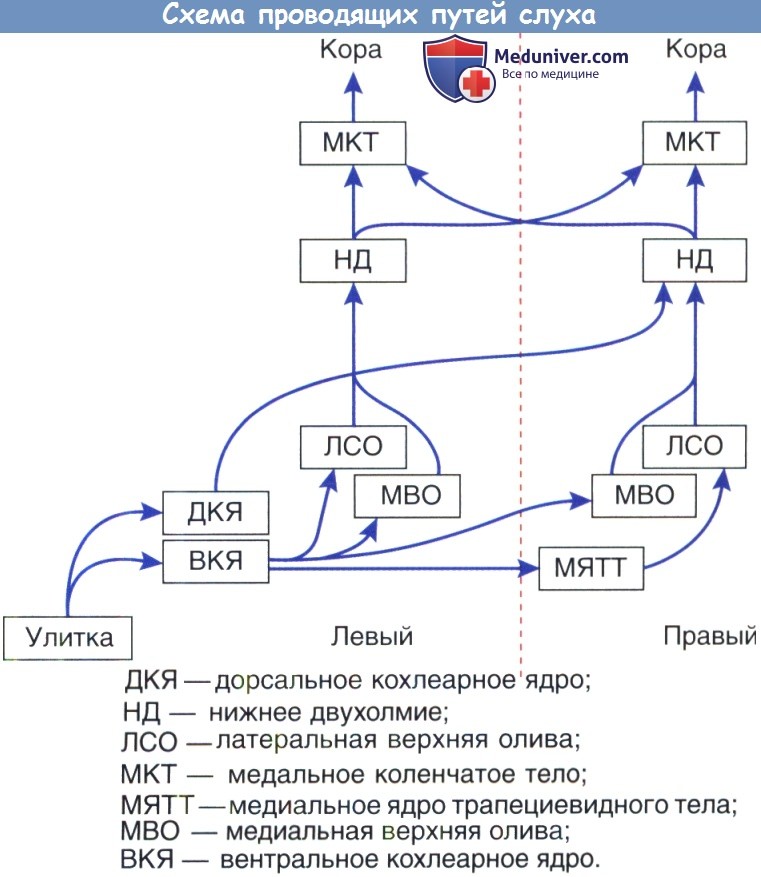
ДКЯ —дорсальное кохлеарное ядро; НД — нижнее двухолмие; ЛСО — латеральная верхняя олива;
МКТ — медальное коленчатое тело; МЯТТ — медиальное ядро трапециевидного тела;
МВО — медиальная верхняя олива; ВКЯ — вентральное кохлеарное ядро.
б) Основные физиологические свойства вентрального кохлеарного ядра (ВКЯ). Анатомическое распределение определенных нейронных групп позволило физиологам связывать анатомическую специализацию нейронов вентрального кохлеарного ядра (ВКЯ) с их свойствами кодирования звука. Ответы в переднем ядре наиболее вероятно регистрируются от мшистых клеток, в то время как реакции в заднем ядре зарегистрируются от «осьминогообразных» клеток. Эти предположения были подтверждены внутриклеточными экспериментами, которые охарактеризовали физиологические свойства нейронов кохлеарного ядра перед маркировкой для последующей визуализации клетки материалом, таким как пероксидаза корней хрена (ПКХ).
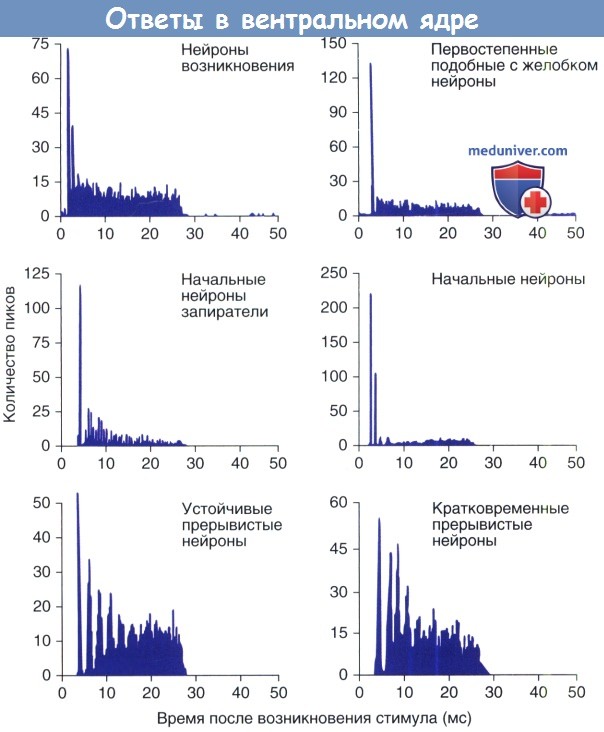
Единичная электрофизиологическая активность в вентральном кохлеарном ядре (ВКЯ), как правило, определяется ответами на короткие импульсы тона на лучшей частоте (наиболее чувствительная частота). Временные характеристики вызванных звуком потенциалов действия или пики регистрируются и суммируются в височной доле из множества стимулов и образуют перистимульную гистограмму (ПСТГ). В зависимости от формы временной гистограммы, могут быть выделены шесть основных видов ответов.
Первичные нейроны дают гистограмму с отражением относительно выраженного ответа, но быстро адаптирующегося к более низкой устойчивой частоте. Так как первичные нейроны обнаруживаются в передних отделах ВКЯ, они объединяются для функционирования подобно кустистым клеткам. В соответствии с высокой стабильностью концевых утолщений, которые связывают кустистые клетки с волокнами слухового нерва, физиологические характеристики первичных нейронов тесно связаны с их периферическими выходами.
Первичные нейроны с желобком обнаружены в областях, содержащих большие количества глобулярных мшистых клеток. Их перистимульная гистограмма содержат серию острых пиков, которая следует за кратким периодом неактивности. Этот «желобок» отражает рефракционный период нейрона после возникновения потенциала действия. Первично подобные нейроны с вырезкой обладают этим свойством, потому что измененные концевые окончания синхронизируют их ответы на начало стимула.
«Начальные» нейроны названы таким образом из-за своего свойства возбуждаться в начале стимула и затем показывать низкую активность к моменту стихания стимула. Начальные нейроны-запиратели (начальные-3), как правило, регистрируются в задних отделах ВКЯ, их морфологическая связь с «осьминогообразными» клетками была установлена посредством внутриклеточной маркировки. Точное возникновение ответа, широкая настройка частоты и широкий динамический диапазон начальных нейронов согласуются с высокой конвергенцией афферентных входов «осьминогообразных» клеток.
Начальные нейроны-прерыватели (начальные-П) менее вариабельны, чем начальные-3 нейроны. Поскольку их потенциалы действия возникают с регулярными интервалами, начальные-П нейроны дают «прерывистую» ПСТГ. Эта реакция приписывается D-звездчатым нейронам, которые являются большими мультиполярными клетками, посылающими дорзальные двусторонние проекции в вентральном кохлеарном ядре (ВКЯ). Эти нейроны также называют «лучистыми» нейронами т.к. их дендритные поля располагаются в трех измерениях, объединяющих входы слухового нерва через широкий диапазон частот. Оказывается, они имеют один источник глицинэргического торможения, который играет важную роль в кодирующих свойствах звука в кохлеарном ядре.
Клетки-прерыватели — физиологическая копия Т-звездчатых клеток. Эти маленькие мультиполярные клетки распространяются вентрально через трапециевидное тело к контралатеральным нижним бугоркам. Эти нейроны также описываются как «плоские» нейроны, так как их дендритные поля ориентированы в плоскости частотой пластины. В дополнение к острой настройке на частоту, прерывистые элементы отражают крайне регулярную частоту выброса и поэтому производят «прерывистый» ПСТГ. Объединяя ответы многих слуховых нервных волокон, клетки-прерыватели производят сигналы, которые устойчивы к изменению уровня звука и эффектам фонового шума.
Типы физиологических ответов в вентральном кохлеарном ядре.
Перистимулирующая гистограмма для 30-милисекундного тонового сигнала.
в) Кодирование звука в вентральном кохлеарном ядре. Разнообразные физиологические структуры групп клеток в вентральном кохлеарном ядре (ВКЯ) наделяют нейроны кодирующими способностями с избирательным подходом к акустическим особенностям сложных звуков. Кроме «начальных» нейронов, проекционные нейроны имеют тенденцию к четкой настройке на частоту. Следовательно, в слуховом нерве сложные спектры с многообразными частотными компонентами должны быть закодированы исходящими частотами множества нейронов с дополнительными возможностями настройки.
С точки зрения человеческого слухового опыта обработка сложных спектральных форм — это основа речевого восприятия.
В английском языке самым элементарным спектром является устойчивый гласный. Перцептивное распознавание гласного звука определяется частотами его формант, макроэргически связанными со стимулами. Эти несущие информацию элементы закодированы сотнями нейронов в слуховом нерве и кохлеарном ядре, объединяющиеся для создания удивительно ясной картины, в которой исходящая частота линейно связана с количеством спектральной энергии в пределах частотного диапазона каждого нейрона.
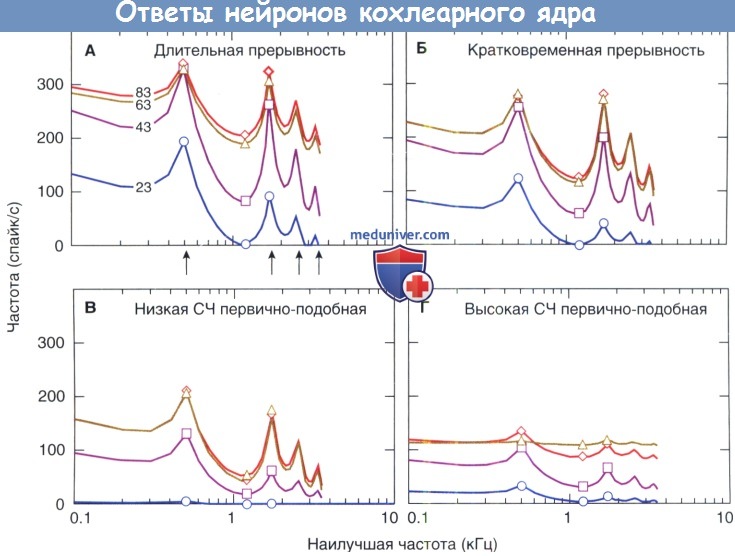
Кривая гласного звука появляется, когда активность, вызванная гласным звуком, отражается на наилучших частотах (НЧ) нейрона. Например, английский гласный /е/ (как русское /э/), как в слове «bet», вызывает высокочастотные выходные сигналы около 0,5 и 1,7 Гц, которые частотно локализуются у его первой и второй формант.
Длительные и кратковременные единицы прерывности обеспечивают превосходные уровни представления спектров гласного звука в широком диапазоне уровней звука. Предполагается, что этот расширенный динамический диапазон обеспечивается плотным схождением слуховых нервных волокон на многополюсных клетках.
Волокна с высокими спонтанными частотами (СЧ) зачастую имеют низкие пороги чувствительности. Единицы прерывности могут ослаблять эти входы на низких уровнях звука. С другой стороны, волокна с низкими СЧ имеют высокие пороги чувствительности, которые могут доминировать над прерывными ответами на высоких уровнях звука. Были зафиксированы обходные пути данного «избирательного слушания», при котором высокочастотные входы локализуются на дистальных дендритах, а низкочастотные входы располагаются около тел прерывающих клеток.
При звуках высокой интенсивности активация промежуточных тормозящих входов из радиальных нейронов отводит насыщенные высокочастотные импульсы от тел клеток. Ответный механизм для низкочастотных импульсов не требуется, так как их действие ограничено порогами чувствительности при низких уровнях шума.
Изображение уровня гласного /e/ (русский /э/) нейронами кохлеарного ядра. Частоты формант указаны стрелками на рис А.
Символы указывают на средние значения ответов нейронов в трех спектральных областях, которые определяют структуру форманты.
Интерполированные точки являются производными линейной закономерности между уровнем их возможностей и исходящей частотой, при которой они возбуждаются.
Числовые обозначения отражают уровень гласного раздражителя.
СЧ — спонтанные частоты.
Репрезентация гласных звуков первично-подобными нейронами ВКЯ демонстрирует ограниченность динамического диапазона входов их слуховых нервов. Первично-подобные нейроны с низкой СЧ обеспечивают скудное представление о формантной структуре на низких уровнях звука, потому что они неэффективно управляются стимулом. Структура гласных звуков проявляется на надпороговом уровне, так как наивысшая исходящая частота ограничена формант-настраивающими нейронами. Первично-подобные нейроны с высокой СЧ обеспечивают хорошую репрезентацию формантной структуры на низких уровнях, но плохой профиль на высоких из-за того, что полный набор нейронов реагирует на максимальной частоте.
Пороги и эффекты насыщения первично-подобных нейронов ограничивают их способность к кодированию спектральной формы (исходящей частоты), однако их ответы богаты временной информацией. В частности, временная точность потенциалов действия является критичной для обобщения пространственной локализации слуховых стимулов.
Большая часть естественных звуков представляют собой краткие переходные процессы. Исходные нейроны активно отвечают на эти стимулы и могут участвовать в тесно-связанных сенсорно-двигательных проводящих путях, которые контролируют реакцию на внезапный акустический стимул и ответные движения. Бинауральное сравнение времени начала стимулов также оказывает сильное влияние на пространственное восприятие. Результат не обязательно должен возникать с началом стимула. Резкий акустический переход стимула по огибающей вызовет подобную реакцию.
Если переходные процессы повторяются с постоянной частотой, то реакции исходных нейронов будут определяться периодом стимуляции. Периодичность — фундаментальная характеристика уровня восприятия и важнейший сигнал для выделения звуковых сигналов из фонового шума.
г) Симптомы и клиника патологии вентрального кохлеарного ядра (ВКЯ). Анализ неврологических расстройств человека был важен для объяснения физиологических основ более высоких когнитивных функций. Текущее понимание локализации центров речи и понимания языка в коре головного мозга можно проследить вплоть до описанных Broca и Wernicke в XIX веке пациентов, страдающих афазией. Более широкое определение клинических проявлений проводящих путей, несущих информацию к коре, было получено при хирургических манипуляциях на экспериментальных животных.
Функциональные нарушения ВКЯ были исследованы путем повреждения путей, которые выходят из ядра и проходят через трапециевидное тело. Итоговые отклонения были по тяжести сопоставимы с последствиями удаления улитки или слухового нерва. Если повреждение трапециевидного тела выполнялось в передне-среднем отделе ствола мозга, то устранялись сигналы от обоих кохлеарных ядер и субъект оказывался глухим. При простом латерализованном повреждении терялся сигнал от одного ВКЯ. Субъекты поддерживали слуховую функцию в одном ухе, но они претерпевали серьезную потерю направленного слушания, потому что мозг больше не проводил бинаурального сравнения источника стимулов.
Временное выключение входящих в ВКЯ стимулов может быть причиной долговременных клинических проявлений. На ранних стадиях развития для установления связей с улитковым ядром требовался звук, активирующий слуховой нерв. Если ухо выключено из-за кондуктивной потери слуха или сенсоневральных нарушений, концевые окончания Хелда подвергаются гипертрофии, которая ассоциируется с временным дефицитом обработки. Синапсы и, вероятно, высшие когнитивные функции, могут быть сохранены срочными вмешательствами, направленными на восстановление поступления входных сигналов к кохлеарному ядру. Эти данные, полученные на моделях животных, могут объяснить, почему кохлеарная имплантация и установка слухового аппарата наиболее эффективны в молодом возрасте.
Схема восходящих путей центральной слуховой системы.
Основные связи между главными ядрами показаны для левого уха.
Симметричная проекция для правого уха не показана.
Учебное видео проводящих путей слухового анализатора
Скачать данное видео и просмотреть с другого видеохостинга можно на странице: Здесь.