Анатомия и физиология дорсального кохлеарного ядра (ДКЯ)
Дорсальное кохлеарное ядро (ДКЯ) является центром мультисенсорной интеграции.37 Основные выходные нейроны биполярны и имеют булавовидные окончания на базальных и апикальных ветвях. Возбуждающие входы от улитки идут к базальным дендритам. Смешанные источники возбуждения и торможения связываются с апикальными дендритами и телами нейронов, поступая от вестибулярного, мозжечкового и соматосенсорного ядер.
Поскольку эти последние входы направляются к голове, шее и наружному уху, предполагается, что мозжечковая схема дорсального кохлеарного ядра (ДКЯ) может быть вовлечена в акустико-моторные поведенческие реакции и локализацию источника звука с основой в ушной раковине.
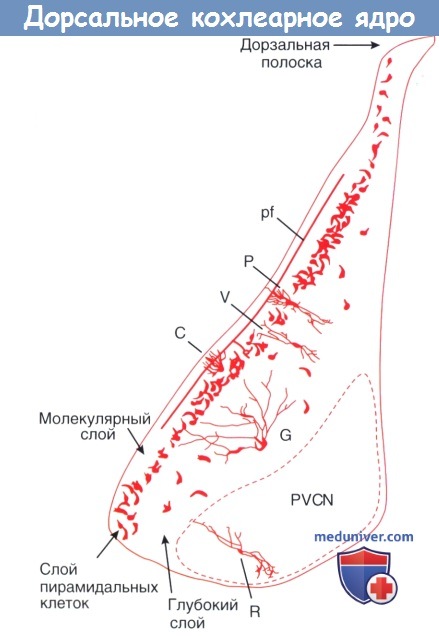
а) Анатомия дорсального кохлеарного ядра (ДКЯ). У большинства млекопитающих ДКЯ является трехслойной структурой. Наружный молекулярный слой содержит нисходящие входы, поступающих в ядро извне и местные целевые нейроны. Разнообразные источники этих «мшистых волокон» были определены при введении ПКХ в дорсальную акустическую полоску (ДАП), где волокна входят в ядро.
После этой процедуры, наблюдалось массивное ретроградное накапливание в комплексе верхней оливы, так как ПКХ подхватывался коллатеральными аксонами оливокохлеарного комплекса. Обширная маркировка неслуховых групп клеток была отмечена в мозжечке и вестибулярных ядрах. Ретроградное накапливание также было обнаружено в боковых клиновидных и сенсорных тройничных ядрах. В совокупности эти структуры несут проприоцептивную информацию от верхней части тела, лица и уха.
Мультисенсорная информация, доставленная к ДКЯ по мшистым волокнам, передается к клеткам-гранулам. Клетки-гранулы — это мелкие клетки, которые сосредоточены вдоль молекулярного слоя и в границах, которые разделяют главные части ядерного комплекса. Аксоны клеток-гранул формируют множество параллельных волокон, которые пересекают ядро на всем протяжении. Эти волокна возбуждают пирамидальные клетки посредством прямых связей с апикальными дендритами или тормозят их через местные интернейроны -колесообразные клетки.
Слой пирамидальных клеток образован крупными проецирующими нейронами ДКЯ. Пирамидальные клетки, известные как веретенообразные клетки, часто располагаются вдоль продольной оси ядра. Они посылают свои апикальные дендриты в скопление клеток молекулярного слоя, где с ними связываются клетки-гранулы и колесообразные клетки. Они направляют базальные дендриты в глубокий слой, где они встречают входы от слуховых нервных волокон и дополнительных источников торможения. Тонотопическая карта улитки передается к ДКЯ систематической иннервацией дорсальной порции слухового нерва.
Глубокий слой ДКЯ заполнен гигантскими и вертикальными клетками. Гигантские клетки — менее часто встречаемые проекционные нейроны. Основные физиологические свойства подобны пирамидальным клеткам, но их глубоко расположенные дендриты не достигают молекулярного слоя и не связываются с параллельными волокнами. Вертикальные клетки являются тормозящими интернейронами. Их узкополосное инг ибирующее влияние отражается расположением вдоль частотной полоски ДКЯ. Дополнительное широкополосное глицеринергическое торможение связано с лучистыми клетками вентрального кохлеарного ядра (ВКЯ).
Анатомическое распределение первичных морфологических классов в рамках слоистой структуры дорсального кохлеарного ядра.
С — веретенообразная клетка; G — клетка-гранула; Р — пирамидная клетка; pf — параллельные волокна;
PVCN — задневентральное кохлеарное ядро; R — лучистая клетка; V — вертикальная клетка.
б) Нейронное кодирование спектральных сигналов для звуковой локализации. Основные физиологические свойства проекционных нейронов дорсального кохлеарного ядра (ДКЯ) формируются сложными взаимодействиями их многочисленных возбуждающих и тормозящих входных сигналов. Изображения ВКЯ линейные, узко настроенные и рассредоточенные, изображения ДКЯ нелинейные, широко настроенные и интегративные. Поэтому показатели разряда индивидуальных пирамидальных и гигантских клеток идеально подходят для кодирования спектральной формы сложных звуков.
Кошка, являющаяся наиболее изученной животной моделью обработки в дорсальном кохлеарном ядре (ДКЯ), соответствующую спектральную информацию получает посредством передаточной функции слухового аппарата (ПФСА).
Эта форма фильтра описывает направленные ответы головы и наружного уха на свободные стимулы, при их распространении к барабанной перепонке. Основанные на ПФСА спектральные сигналы имеют решающее значение для точной локализации высоты источника звука.
Характерной особенностью ПФСА кошки является единственный глубокий спектральный зубец, который резко уменьшается на частотах между 5 и 20 кГц. Когда широкополосной звук проходит через передаточную функцию, эти частоты по существу, отфильтровываются из стимула. Частотная локализация зубца систематически меняется при движении источника звука по высоте или по азимуту. Когда эти сигналы удаляются из свободного звукового поля, то кошки демонстрируют слабонаправленные ориентационные ответы.
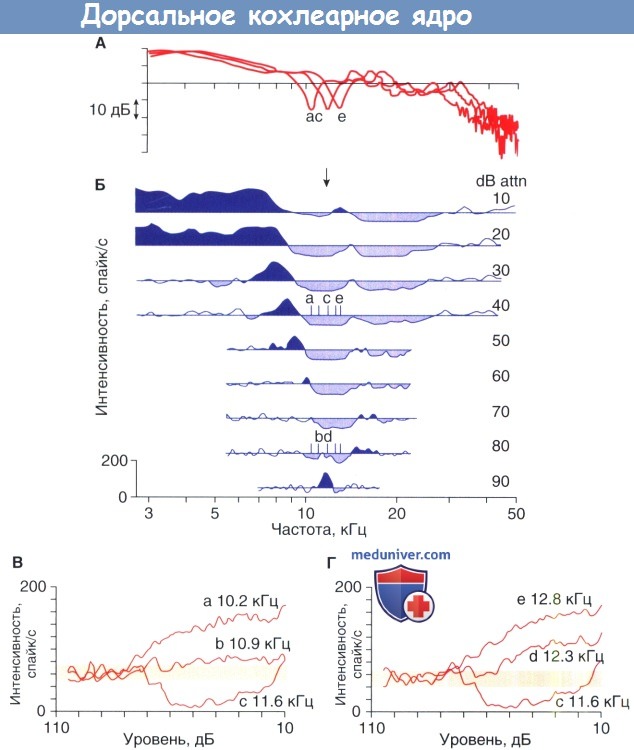
Нейронное кодирование спектральных сигналов для локализации звука было исследовано при записи ПФСА-у-правляемых показателей разряда проекционных нейронов дорсального кохлеарного ядра (ДКЯ), которые известны как IV тип нейронов. Для улучшения контроля стимула эти эксперименты с моделируемыми спектральными зубцами выполнялись с помощью наушников. Зубец может быть смещен по частоте методами обработки цифрового сигнала в соответствии с настройкой нейронов, расположенных вдоль траектории регистрирующего электрода.
Целью этих экспериментов являлась имитация перемещения источника звука, путем манипулирования функции интересов по отношению к ингибирующим свойствам IV типа нейронов. Когда частота зубца ниже оптимальной частоты (ОЧ), нейрон стимулируется исключительно от раздражителя. Аналогичный ответ получают, когда зубец выше ОЧ . Такие результаты ожидаются при учете возбуждающего эффекта широкополосного шума. Однако, когда зубец совпадает с ОЧ, то нейрон в значительной степени ингибируется.
Ответы нейронов IV типа в ПФСА формах диктуются их нелинейными спектральными свойствами интеграции. Следовательно, нейроны избирательно реагируют на спектральные формы вне зависимости от величины индивидуальных частотных компонентов. Определяемое характеристиками, устойчивое представление показано благодаря изменению уровня представления ПФСА. В отличие от более распространенных механизмов кодирование в ВКЯ, основные изменения в спектральной энергии мало влияют на полярность ответов IV типа нейронов.
Спектральное кодирование в дорсальном кохлеарном ядре.
А - Общие передаточные функции головы кошки. Биологически значимый спектральный зубец сдвинут по частоте с изменением частоты дискретизации при воспроизведении.
Б — Отображение частотных ответов IV типа нейронов в дорсальном кохлеарном ядре (ДКЯ).
Буквы указывают на расположение частоты спектрального зубца в пяти частотах дискретизации на оптимальной частоте (ОЧ) нейрона.
Возбуждающие ответы — участки темно-синего цвета. Тормозные ответы — участки светло-голубого цвета.
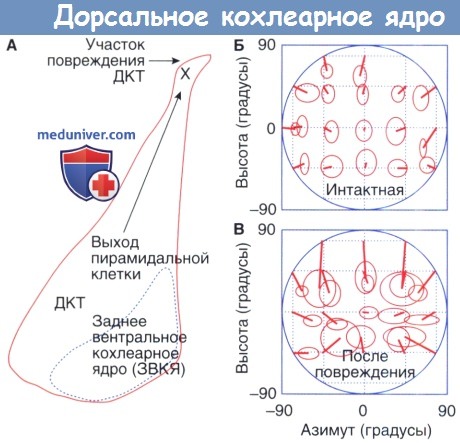
в) Симптомы и клиника поражения дорсального кохлеарного ядра (ДКЯ). Перцептивные нарушения, связанные с нарушениями обработки в ДКЯ, были исследованы в лаборатории путем пересечения дорсального кохлеарного тракта (ДКТ). Эта хирургическая манипуляция устраняет восходящую аксональную инневрацию пирамидальными и гигантскими клетками нижнего бугорка. В отличие от поражений трапециевидного тела, поражения ДКТ мало влияют на основные признаки слуха. Абсолютные пороги в тишине и на фоне шума не зависят от данной процедуры.
Значительные нарушения наблюдаются в тот момент, когда кошек с поражением дорсального кохлеарного тракта (ДКТ) тестируют на локализацию парадигм, требующих идентификации высоты источника звука. Кошки делают рефлекторные движения головой по направлению к внезапным, неожиданным звукам. Они также могут быть обучены согласованному выполнению данных движений при поощрении пищей. Кошка хищник по своей природе, и точность этих управляемых звуком реакций приближается к границам человеческого направленного слуха.
Когда выходные проводящие пути дорсального кохлеарного ядра (ДКЯ) с двух сторон разрушены, кошки демонстрируют явные проблемы с ориентацией. Несмотря на то, что выполнение находится под угрозой и в горизонтальном, и в вертикальном направлениях, наибольшие ошибки наблюдаются в определении высоты источника звука. Эти систематические ошибки подтверждают, что ДКЯ является важным участком для слуховой обработки спектральных сигналов с целью определения локализации звука.
Кошки с двусторонними поражениями дорсального кохлеарного тракта (ДКТ) сохраняют направленный слух в горизонтальном направлении путем участия бинауральных сигналов, которые обрабатываются в вентральных путях, ведущих к комплексу верхней оливы.
У людей редко встречается полное торможение выходов ДКЯ при травме сосудов. Тиннитус (шум в ушах) может отражать не полностью картину разрушения этой деликатной структуры. Если текущие физиологические обоснования шума в ушах подтвердятся, то нарушения обработки в ДКЯ отражают наиболее частую форму нарушений слуха в современном индустриальном обществе.
Эффекты от повреждения дорсального кохлеарного тракта (ДКТ) на слуховую ориентацию.
А — место поражения. Б — точность слуховой ориентации у интактной кошки.
В — Точность после повреждения дорсального кохлеарного тракта (ДКТ) с обеих сторон.
Эллипсы указывают стандартное отклонение реакций в горизонтальных и вертикальных измерениях.
Существует достаточно доказательств того, что частота спонтанных разрядов в ДКЯ увеличивается, когда кохлеарные входы уменьшаются вследствие кратковременного воздействия интенсивного звука или долгосрочных процессов старения. Поскольку эта гиперактивность имитирует нормальную слуховую реакцию на физический стимул, это создает впечатление фантомного звука.
Функциональные изменения, сопровождающие звон в ушах, были тщательно изучены на моделях животных, позволяя контролировать индукцию расстройства и непосредственную физиологическую оценку последствий. Этот анализ пока не раскрывает однозначный участок источника и редкую патологию, но становится ясно, что потенциальные структуры имеют общие свойства. ДКЯ, НК и слуховая кора связаны с тиннитусом.
Эти центры обработки являются ключевыми участками мультисенсорной интеграции, где входы от слуховых и неслуховых ядер сходятся для того, чтобы действовать в совокупности возбуждающих и тормозных нейромедиаторов. Нарушения любого компонента в этой синаптической конструкции способны перестроить состояние покоя и звукопроводящие свойства нейронной цепи.
Нейронная гиперактивность остается определяющей физиологической моделью тиннитуса. Широкая поддержка этого гипотетического механизма отчасти связана с интуитивно понятной простотой. Кроме того, глобальные изменения в мозговой активности предсказывают метаболическую причину, которая может быть обнаружена неинвазивно у пациентов с шумом в ушах.
Несмотря на то, что некоторые модели на животных реагируют на акустическое чрезмерное воздействие при помощи генерализованной гиперактивности, важные детали предложенной связи между активностью головного мозга и восприятием звука остаются неразрешенными. Результаты могут быть специфичными для видов, способов индукции и методов электрофизиологической записи. Более полное объяснение основных механизмов звона в ушах может быть получено из более сложного статистического анализа моделей регулярности, синхронизации, либо долгосрочных колебаний спонтанной активности. Эти свойства не могут быть адекватно описаны без прямых электрофизиологических записей на моделях животных.